



Метформин: новые горизонты применения
Бигуанид метформин – наиболее широко используемый при сахарном диабете (СД) 2 типа препарат, его принимают около 150 млн пациентов во всем мире. Метформин был получен из Galega officinalis (галега лекарственная, козья рута, французская сирень), традиционного лекарственного растения, широко используемого в средневековой Европе для облегчения симптомов диабета. В конце XIX в. было установлено, что G. officinalis содержит большое количество гуанидина – соединения, у которого впоследствии были обнаружены гипогликемические свойства. Однако клиническому применению гуанидина препятствовала его выраженная токсичность, и исследователи переключили свой фокус на более безопасные аналоги. Бигуаниды, состоящие из двух N-связанных гуанидинов, были синтезированы в 1920-х гг., однако их терапевтический потенциал остался без надлежащего внимания вследствие появления в том же десятилетии инсулинотерапии. И только в 1957 г., после публикации успешного исследования французского врача Жана Стерна, метформин начал использоваться для лечения диабета. В отличие от ранее применявшихся противодиабетических средств преимуществом бигуандидов была способность снижать уровни глюкозы в крови без риска развития гипогликемии. Первоначально популярными были более сильные бигуаниды фенформин и буформин, однако после большого количества сообщений об ассоциированном лактатацидозе эти препараты в 1970-х гг. были отозваны с рынка в большинстве стран. В итоге благодаря хорошему профилю безопасности метформин стал препаратом первой линии терапии СД 2 типа и сегодня входит в перечень жизненно важных лекарств Всемирной организации здравоохранения. В настоящее время очевидно, что терапевтический потенциал метформина выходит далеко за рамки лечения диабета. Появляется все больше данных, демонстрирующих роль препарата в терапии множества заболеваний, включая рак и кардиоваскулярную патологию. Кроме того, имеются доказательства того, что метформин замедляет процесс старения и модулирует микробиоту, которая, как известно, имеет огромное значение для поддержания здоровья человека. В настоящем обзоре обсуждаются предложенные молекулярные механизмы, объясняющие эти разнообразные эффекты.
Метформин и СД 2 типа
Согласно данным статистики, в мире в настоящее время проживает примерно 382 млн пациентов с СД. Из-за урбанизации и связанного с ней роста распространенности ожирения и малоподвижного образа жизни в 2035 г. эта цифра, по прогнозам, достигнет 592 млн. СД 2 типа, на который приходится 85-95% случаев диабета, характеризуется гипергликемией, возникающей вследствие инсулинорезистентности или нарушенной секреции инсулина. Это заболевание с комплексной этиологией, включающей взаимодействие между множеством генетических и внешних факторов. Сильными предикторами СД 2 типа являются отягощенный семейный анамнез, повышенный индекс массы тела, высокое артериальное давление, низкая физическая активность, нездоровое питание и старший возраст. В долгосрочной перспективе СД 2 типа может приводить к тяжелым и опасным для жизни осложнениям, таким как кардиоваскулярные заболевания, нейропатия, ретинопатия и нефропатия. Развитие этих осложнений можно предотвратить или значительно отсрочить посредством эффективного контроля уровня глюкозы в крови, для достижения которого необходима модификация образа жизни и, во многих случаях, назначение пероральных антигипергликемических препаратов, таких как метформин.
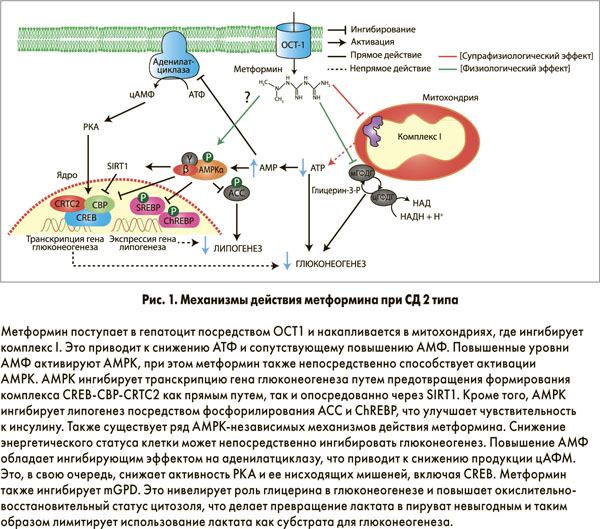
Несмотря на то что метформин применяется в лечении диабета с 1957 г., в настоящее время открываются все новые и новые механизмы его действия. Раньше антигипергликемический эффект препарата связывали с повышением чувствительности гепатоцитов к инсулину и повышенным захватом глюкозы периферическими тканями. Сегодня установлено, что метформин действует преимущественно путем супрессии глюконеогенеза в печени. У пациентов с диабетом метформин может снижать глюконеогенез на 36%. Тем не менее молекулярные механизмы, ответственные за снижение продукции глюкозы, остаются предметом обсуждений. Предложенные механизмы действия метформина при СД 2 типа представлены на рисунке 1.
Молекулярные мишени метформина
Благодаря необычной гидрофильной природе метформин не может пассивно проникать через клеточные мембраны; его поступление в гепатоциты осуществляют органические катионные транспортеры (OCT). Установлено, что терапевтическая эффективность метформина обеспечивается транспортером ОСТ1, при этом генетический полиморфизм ОСТ1 у человека может обусловливать вариабельный ответ на препарат. После захвата гепатоцитом метформин накапливается в митохондриальном матриксе. Считается, что ключевой мишенью метформина является комплекс I дыхательной цепи митохондрий. Этот вывод был сделан по результатам двух независимых работ, показавших, что метформин селективно ингибирует окисление субстратов комплекса I, но не субстратов комплексов II или IV. Эти данные, изначально полученные на культурах изолированных гепатоцитов крыс, впоследствии были подтверждены на многочисленных клеточных моделях первичных гепатоцитов человека.
Механизм ингибирования метформином комплекса I точно не установлен; предполагается, что повышенный уровень рН митохондриального матрикса превращает метформин в депротонированную форму с высокой аффинностью к ионам меди. Эти медные комплексы могут взаимодействовать с чувствительными окислительно-восстановительными реакциями дыхательной цепи, что согласуется с более ранними наблюдениями о зависимости клеточных эффектов метформина от способности препарата связывать медь. Альтернативный механизм ингибирования комплекса I недавно был описан Bridges и соавт. Исследователи продемонстрировали, что метформин неконкурентно ингибирует восстановление убихинона, вероятно, посредством связывания с интерфейсом гидрофильных и мембранных доменов, а также связывания этого фермента в неактивную конформацию с открытой петлей.
Следствием ингибирования комплекса I метформинов является снижение продукции аденозинтрифосфата (АТФ) с сопутствующим повышением уровней аденозинмонофосфата (АМФ) и аденозиндифосфата (АДФ). Этот сдвиг энергетического метаболизма клетки детектируется главным энергетическим сенсором клеток – AMP-активированной протеинкиназой (AMPK).
AMPK-зависимые механизмы
AMPK – эффективный регулятор энергетического гомеостаза клеток, который активируется путем связывания молекул АДФ или АМФ с сайтом его регуляторной γ-субъединицы. Это позволяет клетке отвечать на сниженный энергетический статус переходом с АТФ-потребляющего анаболического состояния в АТФ-продуцирующее катаболическое состояние.
Ключевая роль AMPK в механизме действия метформина была показана в 2001 г. после публикации известного исследования Zhou и соавт. Ученые установили, что метформин стимулирует активацию AMPK в первичных гепатоцитах крыс, и использовали ингибитор AMPK соединение С, чтобы продемонстрировать необходимость AMPK для ингибирования продукции глюкозы под действием метформина. Впоследствии эти данные были подтверждены Saw и соавт., обнаружившими, что утрата печеночной киназы В1 (LKB1), которая является восходящим активатором AMPK, отвечающим за фосфорилирование ее каталитической α-субъединицы, исключает глюкозоснижающие эффекты метформина у мышей, находящихся на богатой жирами диете.
Было высказано предположение, что сигнальный путь LKB1/AMPK, активирующийся метформином, изменяет программу глюконеогенеза клетки путем ингибирования опосредуемого циклическим аденозинмонофосфатом (цАМФ) ответа коактиватора транскрипции 2 (CRTC2), регулируемого элементсвязывающим белком (CREB), – ключевого регулятора экспрессии гена глюконеогенеза). CRTC2 располагается в ядре клетки, где он соединяется с CREB и обеспечивает повышающую регуляцию коактиватора 1α рецептора γ, активируемого пролифератором пероксисом (PGC‑1α), и его нисходящих целевых генов фосфоенолпируваткарбоксикиназы и глюкозо‑6-фосфата. Деактивация CRTC2 также может достигаться путем AMPK-опосредуемой индукции никотинамидфосфорибозилтрансферазы и сопутствующего повышения печеночного сиртуина 1 (SIRT1). Деацетилирование CRTC2 под действием SIRT1 l делает его уязвимым к убиквитинированию, опосредуемому фотоморфогенезом 1, и дегредации, следствием чего является ингибирование экспрессии гена глюконеогенеза. Также было установлено, что AMPK запускает диссоциацию транскрипционного комплекса CREB-CREB-связующий белок (СВР)-CRTC2 путем фосфорилирования CBP в позиции Ser436.
Значимость статуса AMPK как основного медиатора действия метформина была поставлена под сомнение после публикации результатов работы Foretz и соавт. В этом исследовании было установлено, что метформин ингибирует продукцию глюкозы у трансгенных мышей, у которых отсутствуют печеночные каталитические субъединицы AMPK или LKB1. Авторы высказали предположение, что несоответствие между полученными ими результатами и предыдущими наблюдениями Saw и соавт. может быть обусловлено тем, что в последнем исследовании прямой эффект метформина на продукцию глюкозы в печени не оценивался, а изучалось влияние повторного назначения метформина на уровни глюкозы крови натощак. Следовательно, наблюдения Saw и соавт. в действительности могут быть отражением непрямого эффекта на ось AMPK-LKB1 продукции глюкозы в печени, вероятно, опосредуемого супрессией липогенеза под действием AMPK. Хорошо изученной мишенью AMPK является ацетил-КоА-карбоксилаза (АСС) – скоростьлимитирующий фермент, необходимый для образования малонил-КоА, который, в свою очередь, является предшественником липогенеза и ингибитором β-окисления. У мышей ингибирование АСС под действием AMPK регулирует индуцируемое метформином улучшение чувствительности к инсулину. Кроме того, AMPK может вызывать понижающую регуляцию экспрессии ряда липогенных генов. Таким образом, AMPK может отвечать за долгосрочный терапевтический эффект метформина, достигаемый посредством благоприятной модификации не только углеводного, но и липидного метаболизма, что способствует повышению чувствительности к инсулину.
Большинство исследователей полагают, что метформин активирует AMPK путем повышения соотношения АДФ/АМФ посредством ингибирования митохондриального дыхания, однако существуют и другие гипотезы. Например, метаболические изменения, вызываемые метформином в изолированных клетках скелетных мышц, не согласуются с нарушением продукции энергии в митохондриях; скорее, они отражают прямое ингибирование фермента АМФ-деаминазы, что, в свою очередь, приводит к повышению уровней АМФ и активации AMPK. Также существует возможность того, что AMPK сама по себе является прямой мишенью метформина. В частности, установлено, что метформин в низких концентрациях может повышать образование АМФ-αβγ-трехмерного гетеротрехмерного комплекса в гепатоцитах и in vitro, повышая фосфорилирование каталитической α-субъединицы, хотя потенциальные сайты связывания метформина пока обнаружить не удалось. С другой стороны, в ряде исследований было продемонстрировано, что метформин не является прямым аллостерическим активатором. Таким образом, ингибирование дыхательной цепи с сопутствующим повышением уровней АМФ в настоящее время рассматривается как наиболее вероятный механизм, объясняющий активацию AMPK под действием метформина.
AMPK-независимые механизмы
Учитывая неоднозначность статуса AMPK как ключевого медиатора действия метформина, был предложен ряд AMPK-независимых механизмов. Один из них состоит в том, что ассоциированные изменения клеточного энергетического метаболизма могут непосредственно модулировать продукцию глюкозы. Глюконеогенез – энергетически затратный процесс, в котором для синтеза 1 молекулы глюкозы требуется 6 молекул АТФ. Поскольку терапия метформином приводит к снижению уровней АТФ, гепатоцитам приходится соответствующим образом снижать продукцию глюкозы. Так, Foretz и соавт. установили сильную корреляцию между снижением содержания АТФ и ингибированием продукции глюкозы в первичных мышиных гепатоцитах, инкубируемых с метформином, что свидетельствует о тесной взаимосвязи энергетического статуса гепатоцитов и продукции глюкозы в печени. Кроме того, изменения энергетического статуса клеток, вызываемые метформином, могут подавлять глюконеогенез путем аллостерического ингибирования эссенциальных ферментов. Например, АМФ действует синергически с фруктозо‑2,6-бисфосфатом, ингибируя ключевой фермент глюконеогенеза фруктозо‑1,6-бифосфатазу.
Наличие у метформина молекулярных механизмов действия, не зависящих от транскрипционных изменений, подтверждается в исследованиях по изучению экспрессии генов. Форсированная экспрессия PGC‑1α (главного коактиватора генов глюконеогенеза) не препятствует проявлению метформин-индуцированного снижения продукции глюкозы в гепатоцитах. В ряде исследований также было установлено отсутствие корреляции между экспрессией генов глюконеогенеза и продукцией глюкозы в печени как на экспериментальных моделях у мышей, так и у пациентов с СД 2 типа. В целом результаты этих работ свидетельствуют о том, что ингибирование экспрессии генов глюконеогенеза не является основной детерминантой клинических эффектов метформина; последние также достигаются благодаря изменениям энергетического метаболизма клеток и ассоциированному снижению глюконеогенеза.
Другой AMPK-независимый механизм действия метформина, включающий антагонизм в отношении эффектов глюкагона, был недавно предложен Miller и соавт. В серии экспериментов in vitro и in vivo на первичных гепатоцитах мышей было продемонстрировано, что метформин и еще один бигуанид фенмормин блокируют глюкагон-индуцированную активацию аденилатциклазы, что приводит к снижению синтеза цАМФ. Это, в свою очередь, снижает активность протеинкиназы А (PKA), предотвращая фосфорилирование критических субстратов глюконеогенеза, таких как 6-фосфофрукто‑2-киназа / фруктозо‑2,6-бифосфатаза 1, CREB‑1 и рецептор инозитолтрифосфата. По мнению исследователей, повышение внутриклеточных уровней АМФ, индуцируемое метформином, отвечает за ингибирование аденилатциклазы, вероятно, путем прямого связывания аденинового остатка АМФ с ингибиторным Р-сайтом.
Тем не менее следует отметить, что если антагонизм к глюкагону был бы основным механизмом действия метформина, применение этого препарата ассоциировалось бы с частыми эпизодами гипогликемии, как это наблюдается у трансгенных мышей, не имеющих рецепторов глюкагона. Однако одним из основных преимуществ метформина является очень низкая частота гипогликемии, поэтому его потенциальный антагонизм к глюкагону у человека, по-видимому, не полный или уменьшается компенсаторными механизмами.
Исследования, на которых основаны вышеуказанные модели AMPK-независимых эффектов метформина, подвергались критике за то, что в них использовались концентрации препарата, значительно превышающие максимальные терапевтические у пациентов с диабетом. Метформин назначается перорально, максимальная рекомендованная доза составляет 2,5 г/сут. Было продемонстрировано, что после приема однократный дозы 1,5 г пиковая концентрация в плазме у человека достигает примерно 4 мкг/мл, или 18 мкмоль. В то же время данные, полученные в экспериментах на животных, показали, что концентрация метформина в портальной вене печени значительно превышает таковую в системной плазме.
В исследованиях Foretz и соавт. и Miller и соавт. для демонстрации AMPK-независимых эффектов метформина действительно использовались высокие концентрации препарата, однако это не означает, что указанные эффекты не будут проявляться на клинически значимом уровне при применении более низких концентраций. В экспериментах in vivo на грызунах метформин часто назначается в дозах 250-350 мг/кг. Эти значения были получены с использованием стандартного метода конвертации доз между видами, основанного на нормализации площади поверхности тела. Согласно данной формуле стандартная терапевтическая доза 20 мг/кг у взрослого человека (масса тела 60 кг) эквивалентна дозе примерно 250 мг/кг у мыши. Кроме того, установлено, что у мышей с диабетом для проявления терапевтического эффекта метформин приходится назначать в относительно высоких дозах. Также продемонстрировано, что метформин накапливается в очень высоких концентрациях во многих тканях. У мышей доза 50 мг/кг создает концентрации более 250 мкмоль в печени и еще более высокие концентрации в тонком кишечнике. Таким образом, в исследованиях in vitro на первичных гепатоцитах совершенно оправдано использование концентраций метформина, превышающих таковые в плазме или портальной вене. Кроме того, известно, что биологические эффекты метформина являются время- и концентрационно-зависимыми, что, по-видимому, отражает свойство препарата накапливаться в митохондриальном матриксе. Это означает, что эффекты, наблюдаемые при высоких концентрациях, также могут проявиться и при более низких концентрациях после более продолжительного воздействия метформина.
Исследования, в которых для изучения AMPK-независимых механизмов действия метформина использовались нокаутированные по печеночной AMPK животные, следует интерпретировать с учетом недавно полученных доказательств важной роли тонкокишечной AMPK в реализации эффектов метформина. Главным местом действия метформина считается печень, однако активация AMPK в тонком кишечнике вносит существенный вклад в реализацию быстрого глюкозоснижающего эффекта препарата. В экспериментальном исследовании на инсулинорезистентных крысах было продемонстрировано участие оси «пищеварительный тракт – головной мозг – печень»: под действием метформина рецептор AMPK-глюкагоноподобного пептида‑1 в двенадцатиперстной кишке передает сигналы в центральную нервную систему, которая, в свою очередь, дает «команду» печени снизить продукцию глюкозы. Результаты этого исследования также показали, что активация AMPK в органах, как правило, при более высоких концентрациях метформина (в милимолярном диапазоне), может отдаленно регулировать метаболизм тканей, непосредственно отвечающих за продукцию глюкозы.
Недавно был обнаружен новый механизм действия метформина, не зависящий ни от активации AMKP, ни от изменений энергетического статуса клетки. Madiraju и соавт. установили, что метформин ингибирует фермент глицерофосфатного шаттла митохондриальную глицерофосфатдегидрогеназу (mGPDG). Это предотвращает прямое использование глицерина как субстрата для глюконеогенеза, а также приводит к повышению окислительно-восстановительного статуса цитозоля, что делает невыгодным превращение лактата в пируват и таким образом ограничивает роль лактата в глюконеогенезе. Это исследование подчеркивает значимость метформина как неспецифического препарата, способного влиять на множество молекулярных мишеней с итоговым подавлением глюконеогенеза.
Метформин и кардиоваскулярные заболевания
К кардиоваскулярным заболеваниям (КВЗ) относятся болезни сердца и кровотока. По данным Британского фонда сердца (BHF), в Великобритании КВЗ являются причиной более четверти всех летальных исходов и обусловливают экономические потери в сумме 19 млрд фунтов стерлингов в год. Одним из самых сильных факторов риска развития КВЗ является СД 2 типа. Имеются доказательства, что метформин может защищать от КВЗ пациентов с диабетом. Это особенно важно с учетом того, что КВЗ являются ведущей причиной смерти больных СД. В масштабном исследовании UKPDS лечение метформином снизило риск инфаркта миокарда (ИМ) на 39% по сравнению с традиционной терапией за 10-летний период. Последующие исследования подтвердили благоприятную роль метформина в защите против кардиоваскулярных осложнений СД. Безусловно, эти положительные эффекты метформина частично могут быть обусловлены улучшением метаболизма глюкозы и липидов, однако существует и ряд других механизмов, объясняющих непосредственное влияние препарата на кардиоваскулярную систему.
Результаты многочисленных исследований свидетельствуют о том, что метформин обладает антиатеросклеротическими эффектами за счет улучшения целостности эндотелия и предотвращения формирования атеросклеротических бляшек. В частности, было установлено, что активация AMPK под действием метформина уменьшает повреждение эндотелиоцитов, вызываемое окислительным стрессом на фоне гипергликемии. Данный процесс опосредуется ингибированием сигнального пути протеинкиназы С-NAD(P)Н-оксидазы. В результате снижение продукции активных форм кислорода (АФК) в цитозоле останавливает инициацию петли положительной обратной связи между образованием АФК в митохондриях и расщеплением последних, что, в свою очередь, предотвращает эндотелиальный апоптоз путем снижения мембранного потенциала митохондрий.
В условиях инсулинорезистентности метформин также проявляет антитромботические свойства. Например, метформин противодействует стимулирующему эффекту гиперинсулинемии на продукцию ингибитора активатора плазминогена 1 (PAI‑1) – отрицательного регулятора фибринолиза, принимающего участие в образовании кровяного сгустка. Предполагается, что этот эффект опосредуется не улучшением чувствительности к инсулину, ассоциированным с метформином, а непосредственным ингибированием экспрессии гена PAI‑1. Кроме того, у пациентов с инсулинозависимым диабетом лечение метформином значительно уменьшает агрегацию тромбоцитов. Это наблюдение может объясняться результатами экспериментального исследования, в котором было обнаружено, что метформин AMKP-зависимо усиливает фосфорилирование Ser1179 эндотелиальной синтазы оксида азота (NO), что улучшает биодоступность NO для эндотелия крупных сосудов. Как известно, NO играет ключевую роль в поддержании сосудистого гомеостаза, выполняя множество функций, включая ингибирование агрегации тромбоцитов, и таким образом вносит существенный вклад в защиту от развития ишемической болезни сердца (ИБС).
Помимо эндотелийпротекторных эффектов метформин воздействует на функцию кардиомиоцитов, что в экспериментальных исследованиях ассоциировалось с уменьшением диабетической кардиомиопатии. Последняя является ведущей причиной сердечной недостаточности и характеризуется сосудистой дисфункцией, развивающейся независимо от ИБС и артериальной гипертензии. Высказывалось предположение, что метформин может корригировать нарушения релаксации кардиомиоцитов путем тирозинкиназозависимой модуляции кальция. Впоследствии было продемонстрировано, что уменьшение диабетической кардиомиопатии при лечении метформином ассоциируется с AMPK-опосредованной повышающей регуляцией аутофагии. Кардиальная аутофагия является важным механизмом гомеостаза, подавляющим диабетическую кардиомиопатию. AMPK, активируемая метформином, защищает кардиомиоциты путем разрушения комплекса беклин-1-Blc‑2 (известного также как комплекс В-клеточной лимфомы 2) и переключения процессов апоптоза на аутофагию. В экспериментальных исследованиях на мышах с диабетом восстановление аутофагии улучшало структуру и функцию сердца.
Имеются доказательства, что метформин проявляет кардиопротекторные эффекты в отношении повреждения вследствие ишемии-реперфузии после ИМ. Назначение метформина на протяжении первых 15 мин после реперфузии ограничивало размеры инфаркта в изолированных сердцах крыс с диабетом и без диабета. Этот эффект опосредуется активацией сигнального пути протеинкиназы В (Akt) / фосфатидилинозитол‑4,5-бифосфат‑3-киназы (PI3K), что предотвращает открытие митохондриальной поры переходной проницаемости (mPTP) – ключевого триггера клеточной смерти во время реперфузии. Уменьшение размеров инфаркта при лечении метформином также объясняется AMPK-опосредованным повышением активности эндотелиальной синтазы NO. Интересно, что новый низкомолекулярный активатор AMPK (A‑76922) действует синергически с метформином, что проявляется в усилении кардиопротекторных эффектов активации AMPK.
Метформин может уменьшать повреждение, обусловленное ишемией-реперфузией, не только в остром периоде, но и в долгосрочной перспективе. Так, в экспериментальном исследовании назначение метформина на протяжении 4 нед уменьшало размеры ИМ независимо от наличия диабета. При этом кардиопротекторные эффекты, наблюдавшиеся у животных с диабетом, ассоциировались с улучшением структуры митохондрий, вероятно, за счет активации AMPK и повышения экспрессии PGC‑1α. Длительное лечение метформином после ИМ также может оказывать положительный эффект. Было установлено, что 12-недельная терапия метформином после ИМ уменьшает ремоделирование сердца и таким образом может отсрочивать развитие сердечной недостаточности.
Разнообразные положительные эффекты метформина на кардиоваскулярную систему, в том числе при диабете, в настоящее время изучаются в рандомизированных контролированных исследованиях.
Метформин и рак
Во всем мире злокачественные новообразования являются ведущей причиной смерти. Проведенный недавно в Великобритании анализ показал, что более чем у 50% людей в возрасте до 65 лет в тот или иной период жизни диагностируется рак. Высокая социально-экономическая значимость онкопатологии заставляет расходовать огромные средства на исследования новых методов лечения. Несмотря на это, только 5% противоопухолевых препаратов, достигающих I фазы клинических исследований, в итоге получают одобрение, и даже наиболее современные таргетные препараты лишь незначительно улучшают выживаемость.
Интерес к метформину в области онкологии возник в 2005 г. после публикации эпидемиологического исследования, в котором была установлена связь между лечением этим препаратом и сниженным риском рака у пациентов с диабетом. В последующем в многочисленных наблюдательных исследованиях подтвердилась протекторная роль метформина в отношении различных злокачественных опухолей, включая гепатоцеллюлярную карциному, колоректальный рак, рак желудка и пищевода, в диабетической популяции. Способность метформина изменять метаболизм опухолевых клеток и взаимодействовать с разнообразными метаболическими путями свидетельствует о том, что этот препарат может быть эффективным в предотвращении развития и замедлении прогрессирования различных типов рака. Предполагаемые механизмы противоопухолевых эффектов метформина представлены на рисунке 2.
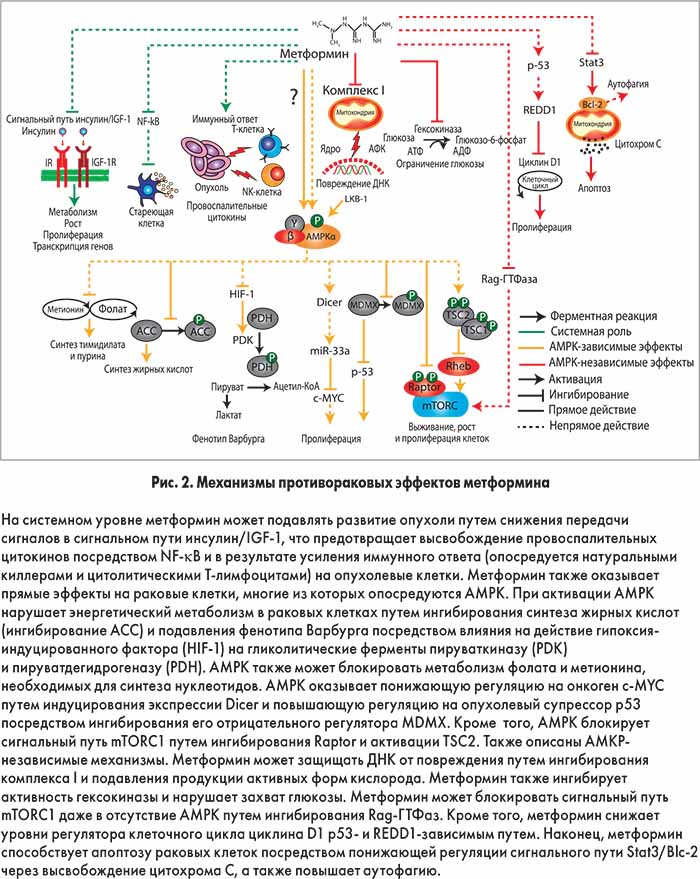
Возможно, защиту от рака обеспечивают системные эффекты метформина. В экспериментальных и эпидемиологических исследованиях было продемонстрировано, что инсулин и инсулиноподобный фактор роста 1 (IGF‑1) могут способствовать развитию злокачественных опухолей за счет стимулирования пролиферации эпителиальных клеток. Метформин может предотвращать непластическую активность путем снижения гиперинсулинемии и уровней этих сигнальных молекул. Кроме того, метформин может модифицировать воспалительные процессы, принимающие участие в прогрессировании рака. Например, установлено, что метформин блокирует активность ядерного фактора транскрипции каппа В (NF-κB), уменьшая секрецию провоспалительных цитокинов стареющими клетками. Этот механизм также объясняет антиэйджинговые свойства препарата. Недавно было продемонстрировано, что метформин усиливает иммунный ответ на раковые клетки, в частности защищает опухоль-инфильтрирующие лимфоциты CD8+ от апоптоза и функционального истощения. Многообещающей находкой стало также то, что метформин способен повышать эффективность экспериментальной противоопухолевой вакцины путем улучшения выживаемости Т-клеток памяти.
Продолжение следует.
СТАТТІ ЗА ТЕМОЮ Ендокринологія
За визначенням Всесвітньої організації охорони здоров’я, цукровий діабет (ЦД) – це група метаболічних розладів, що характеризуються гіперглікемією, яка є наслідком дефектів секреції інсуліну, дії інсуліну або обох цих чинників. За останні 15 років поширеність діабету зросла в усьому світі (Guariguata et al., 2014). Згідно з даними Diabetes Atlas (IDF), глобальна поширеність діабету серед осіб віком 20-79 років становила 10,5% (536,6 млн у 2021 році; очікується, що вона зросте до 12,2% (783,2 млн у 2045 році (Sun et al., 2022). Наразі триває Програма профілактики діабету (ППД), метою якої є визначити, які підходи до зниження інсулінорезистентності (ІР) можуть допомогти в створенні профілактичних заходів ЦД 2 типу (The Diabetes Prevention Program (DPP), 2002). У цьому світлі визначення впливу вітаміну D на розвиток ЦД є актуальним питанням....
Внутрішній біологічний годинник людини тісно та двоспрямовано пов’язаний зі стресовою системою. Критична втрата гармонійного часового порядку на різних рівнях організації може вплинути на фундаментальні властивості нейроендокринної, імунної та вегетативної систем, що спричиняє порушення біоповедінкових адаптаційних механізмів із підвищеною чутливістю до стресу й уразливості. Поєднання декількох хвороб зумовлює двоспрямованість патофізіологічних змін....
Метформін – протидіабетичний препарат першої лінії, який пригнічує глюконеогенез у печінці і в такий спосіб знижує рівні глюкози в крові. Крім того, він знижує ризик кардіоваскулярних подій, чинить нефропротекторний ефект і здатен подовжувати тривалість життя. Завдяки цим властивостям метформін нині розглядають як мультифункціональний препарат і дедалі частіше застосовують для лікування та профілактики різноманітних захворювань....
Сучасне лікування хворих на цукровий діабет (ЦД) 2 типу включає зміну способу життя і медикаментозну терапію для контролю глікемії та профілактики ускладнень. Проте дослідження показують, що на практиці небагато хворих досягають контролю захворювання (частково через погану прихильність до лікування). Частка пацієнтів, які дотримуються протидіабетичної терапії, коливається від 33 до 93% (упродовж 6-24 міс) [1, 2]....