


2 квітня, 2018
Система «теломеры-теломераза» как молекулярно-генетический индикатор старения у человека
Достижения медицинской науки за последние 100 лет почти удвоили продолжительность жизни населения в развитых странах. Так, ожидаемая продолжительность жизни в 2015 г. составляла 71,4 года (73,8 года для женщин и 69,1 года для мужчин). В период между 2000 и 2015 гг. средняя продолжительность жизни увеличилась на 5 лет, что является самым быстрым ростом с 1960-х гг. По статистическим оценкам, этот бум долголетия приведет к тому, что к 2025 г. численность популяции людей в возрасте старше 60 лет достигнет 1,2 млрд.
Резкое старение населения во всем мире и развитие хронических заболеваний с возрастом приводит к значительному вложению средств в программы лечения пожилых людей [1].
Для повышения качества жизни при снижении реальных темпов старения и увеличении продолжительности активной трудоспособности необходима разработка точных количественных методов диагностики процессов, связанных со старением.
Чтобы поддержать здоровье и благополучие указанной популяции, обязательно нужно понимать молекулярные механизмы и биологические процессы, лежащие в основе старения и ассоциированных с ним заболеваний.
Старение представляет собой процесс, вызываемый комплексом регуляторных факторов, и является генетически детерминированным.
Существуют различные подходы к оценке биологического возраста [2]. Последний определяется на основании известных для данной популяции зависимостей между специфическими биомаркерами и хронологическим возрастом по индивидуальным значениям указанных показателей у человека.
Для этого предложены различные тесты (около 600), которые могут служить индикаторами старения. Пока не удается однозначно ответить на вопрос, какое число показателей оптимально для определения биологического возраста.
В настоящее время существуют 2 основные теории механизмов старения: теломерная и окислительная (свободнорадикальная или митохондриальная). Тем не менее появляется все больше доказательств того, что центральная роль в возрастной дегенерации тканей и уменьшении резервов стволовых клеток принадлежит именно теломерам. Ряд исследователей считают теломерную теорию наиболее убедительной.
Теломеры
В 1938 г. американский генетик H. J. Müller, работая в Эдинбургском институте генетики животных (Великобритания), проводил опыты с мухами вида Drosophila melanogaster, подвергая их воздействию рентгеновских лучей [3]. Он заметил, что гены, расположенные на концах облученных хромосом, в отличие от других генов, не претерпевают изменений в виде делеции или инверсии благодаря наличию защитного «колпачка», который он назвал «защитный ген», а позже «теломера» (от греч. telos – конец и meros – часть) (рис. 1).
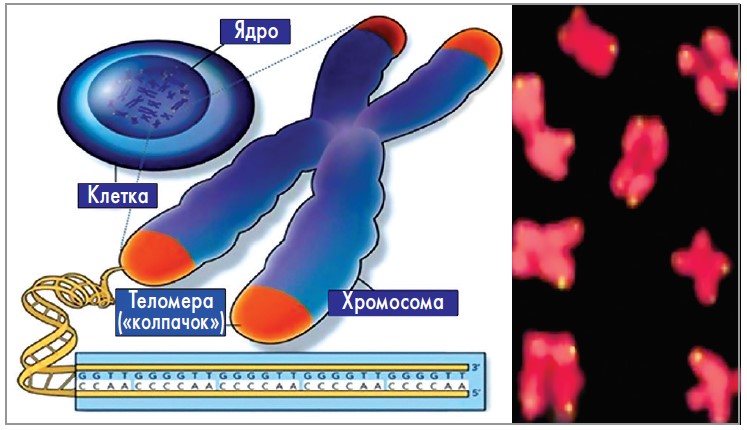
Рис. 1. Схема строения теломеры в составе хромосомы (а)
и митотические хромосомы с теломерами, меченными
флуоресцеином на концах (б)
Два года спустя исследователь из Миссурийского университета (США) B. McClintock [4], работа которой была посвящена изучению генетики кукурузы (Zea mays), обнаружила, что разрыв хромосом приводит к адгезии и слиянию их концов с последующим образованием дицентрических хромосом. Независимо от повреждения, концы могут быть восстановлены благодаря новой теломере. Согласно ее выводам теломеры играют решающую роль в целостности хромосом, поскольку они предотвращают появление циклов «разрывслияниемост», которые катастрофичны для выживаемости клеток.
В 1961 г. L. Hayflick, профессор анатомии Калифорнийского университета (США), и P. Moorhcad [5] представили данные о том, что даже в идеальных условиях культивирования фибробласты эмбриона человека способны делиться только ограниченное число раз (50±10), после чего их способность к пролиферации исчерпывается. Данное свойство, получившее название «клеточное старение» («предел Хейфлика»), наследуется генетически и не зависит от условий культивирования клеток (рис. 2).
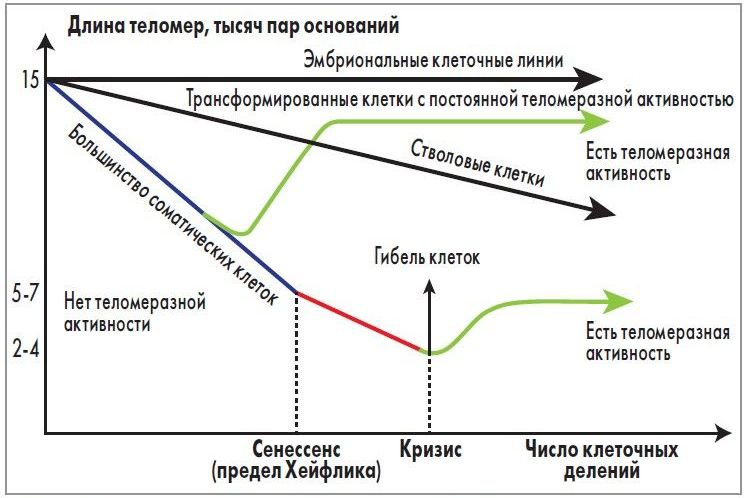
Рис. 2. Связь между длиной теломерной ДНК и числом клеточных
делений
В 1971 г. А. Оловников [7, 8] на основании оригинальных работ J. Watson [6] предложил теорию маргинотомии (отсчета клеточных делений и старения), которая объясняла механизм работы теломер. При матричном синтезе полинуклеотидов ДНК-полимераза не в состоянии полностью воспроизвести линейную матрицу, и реплика получается всегда короче в ее начальной части.
Таким образом, при каждом делении ДНК клетки укорачивается, что ограничивает пролиферативный потенциал клеток и является т. н. счетчиком количества делений и, соответственно, продолжительности жизни клетки. При каждой репликации хромосом теломеры укорачиваются, т. е. дочерняя ДНК оказывается несколько короче материнской (рис. 3).
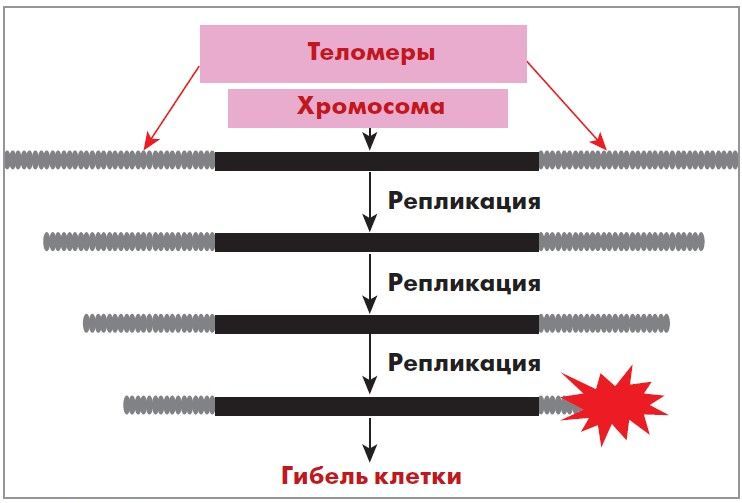
Рис. 3. Укорочение участков теломер в процессе деления
соматической клетки
Теломеры
Теломеры человека – это нуклеопротеидные концевые участки хромосом, содержащие от 4 до 14 тыс. высококонсервативных пар оснований и состоящие из повторов TTAGGG.
Длина участков TTAGGG варьирует от вида к виду. Так, теломеры человека имеют размер 414 kbр (килобазная пара представляет собой единицу длины нуклеиновых кислот, равную 1000 пар оснований), в то время как лабораторные мыши имеют теломеры более длинные – 36150 kbр. Теломера заканчивается однонитевым нависающим 3`-концом, завернутым в структуру, называемую Тпетлей, которая не позволяет концам хромосом соединяться друг с другом. При этом однонитевый конец теломеры проникает в прилегающий к Т-петле двунитевый участок, образуя D-петлю (displacement loop), закрепляющую T-петлю.
Функции теломер зависят от минимальной длины теломерных повторов и активности связанного с ними белкового комплекса.
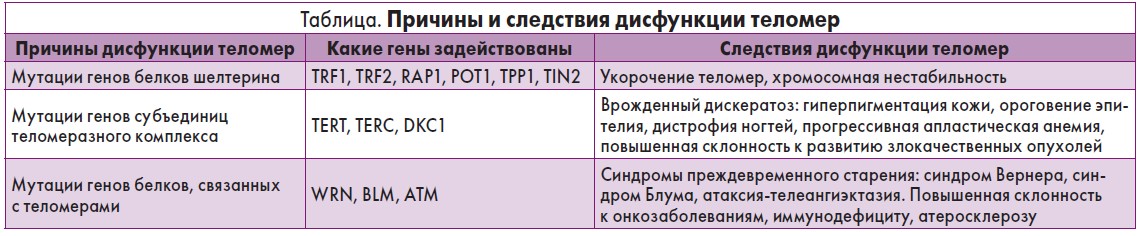
Этот комплекс (шелтерин) состоит из 6 белков [9, 10]:
1. TIN2 (TRF1interacted nuclear protein 2) – препятствует присоединению теломеразы к теломере.
2. TPP1 (Telomere protected protein 1) – образуя гетеродимер с POT1, присоединяется к однонитевой теломерной ДНК, защищая ее от повреждений.
3. POT1 (Protection of telomeres) защищает G-богатую нависающую нить ДНК, располагаясь в олигонуклеотид/олигосахаридном связующем сгибе в теломерах, препятствует слиянию теломер «конецвконец»;
4. TRF2 (Telomere repear binding factor 2) – поддерживает целостность теломер, защищая от слияния друг с другом.
5. TRF1 (Telomere repeat binding factor 1) – отрицательный регулятор длины теломер, не допускающий теломеразу к теломере.
6. RAP1 (Rifassociated protein) – с С-концом этого белка связывается белок (белки) RIF1 и RIF2 (Relaxing insulinlike factors), препятствующие работе теломеразы. Он удерживает Т-петлю, образуя поперечные сшивки с теломерной ДНК.
Функции теломер:
- сохранение целостности генома;
- облегчение гомологичной рекомбинации при мейозе;
- участие в архитектонике клеточного ядра;
- регуляция экспрессии генов (эффект положения гена);
- определение репликативного потенциала клетки.
Хромосомы, лишенные теломер из-за разрывов, подвергаются слиянию, деградации и перестают выполнять свои функции.
Причины и следствия дисфункции теломер представлены в таблице.
Теломераза
В конце 1980-х гг. теорию А. Оловникова подтвердили исследователи из США E. H. Blackburn, J. W. Szostak и C. W. Greider (Нобелевская премия 2009 года с формулировкой «за открытие механизмов защиты хромосом теломерами и фермента теломеразы»).
Им удалось выяснить механизм, благодаря которому хромосомы копируются во время деления клетки и каким образом они защищены от деградации. Учеными было обнаружено, что чем короче теломеры, тем старее клетка, и наоборот, если активность теломеразы, достраивающей теломеры, высокая и постоянно поддерживается одинаковая длина теломер, клетка не стареет.
Теломераза – это большой рибонуклеопротеиновый комплекс, ответственный за синтез ДНК повторов теломер (TTAGGG) на 3`-концах линейных хромосом, что предотвращает потерю ДНК при каждой репликации. Теломераза представляет собой обратную транскриптазу, которая состоит из каталитической белковой субъединицы, называемой обратной транскриптазой теломеразы (TERT) и кодируемой геном hTERT, которая расположена на хромосоме 5p15.33; РНКкомпонента, известного как теломераза РНК человека или теломеразный РНКкомпонент человека (TERC), кодируемого геном hTERC на хромосоме 3q26 и дискерина (кодируется геном DKC1). TERT содержит 1131 аминокислоту и имеет молекулярную массу >100 кД. По своей структуре TERT напоминает форму рукавицы, что позволяет этому ферменту прикрепляться к хромосоме и добавлять в нее участки теломер, состоящие из одной цепочки. TERT захватывает TERC (имеющую длину 451 нуклеотид), которая не транслируется [11].
Теломераза имеет полимеразную активность, при этом она строит цепь ДНК комплементарно своей собственной определенной РНК-матрице.
Последняя, как и вся РНК в клетке, транскрибируется со своего гена, претерпевает изменения, созревает, узнается теломеразой и образует с ней комплекс, который распознает теломерные участки ДНК и удлиняет 3`-конец ДНК на 8 нуклеотидов, комплементарных РНКматрице.
Затем она пересаживается на только что синтезированный 3`-конец и удлиняет его снова. Таких удлинений может быть много, поэтому теломерные участки состоят из множественных повторов.
После этого на удлиненную цепь отжигается праймер и ДНК-полимераза комплементарно достраивает короткую цепь ДНК.
При матричном синтезе полинуклеотидов теломераза не в состоянии полностью воспроизвести линейную матрицу, реплика получается всегда короче в ее начальной части. В связи с этим после определенного количества делений клетка больше делиться не может. Данный феномен получил название «концевой недорепликации ДНК». Из этого следует важный вывод: возраст человека связан с длиной теломер. Чем старше человек, тем меньше средняя длина его теломер: она варьирует от 15 тыс. нуклеотидных пар (НП) при рождении до 5 тыс. НП при хронических заболеваниях. Максимальная длина теломер – у 18- месячных детей, затем она быстро уменьшается до 12 тыс. НП к 5-летнему возрасту. После этого скорость укорачивания снижается.
Метод измерения теломер Длина теломер клеток человека, в частности лейкоцитов, наиболее часто используется в качестве биомаркера старения [12, 13]. Для измерения их длины применяется метод FISH (Fluorescence In Situ Hybridization), или флюоресценция на месте гибридизации, – цитогенетический метод, используемый для детекции и локализации специфических последовательностей ДНК на хромосомах, мРНК и др. В основе методики лежит гибридизация флуоресцентно меченого ДНК/РНК зонда с комплементарной последовательностью ДНК/РНК. Выявление метки происходит с помощью флуоресцентного микроскопа.
Одним из вариантов FISH является QFISH (Quantitative FISH), разработанный U. Lansdorp и соавт. [14]. QFISH – это количественный метод, который применяется для работы с проточной цитометрией. Изначально использовался для измерения длины хромосом (разрешение 200 bp) путем подсчета числа теломерных повторов. На данный момент QFISH является важным инструментом в изучении роли теломер в процессах старения.
Используя метод QFISH и проточную цитометрию, можно измерить общее количество флуоресценции в клетке, чтобы оценить среднюю длину теломер в хромосомах.
Наиболее быстрое снижение длины теломер происходит в течение первых нескольких лет жизни, но продолжается до конца полового созревания. У взрослых скорость укорочения теломер замедляется. Это более заметно в гранулоцитах, что отражает деление клеток в кроветворных стволовых клетках по сравнению с лимфоцитами.
Теломеразная теория старения Факт о том, что теломеры человека укорачиваются с возрастом, стал известен более 20 лет назад. Люди с короткими теломерами входят в группу риска преждевременной смертности. R. Cawthon и соавт. [15] продемонстрировали, что для 25% 60-летних участников исследования, имевших наиболее длинные теломеры, был характерен в 2 раза более низкий риск смерти, чем для 25% участников с наиболее короткими теломерами. R. Cawthon имел доступ к уникальной базе данных, содержащей законсервированные образцы крови 20-летней давности. В течение многих лет никому не удалось ни воспроизвести, ни опровергнуть эти результаты.
Исследования молекулярных механизмов старения осложняются тем фактом, что последнее не является уникальным клеточным процессом. Механизмы старения существенно отличаются у клеток, полученных от различных видов животных.
Мутации в генах, кодирующих фермент теломеразу, включая TERT, TERC и DKCI, приводят к таким заболеваниям, как врожденный дискератоз, апластическая анемия, легочный фиброз, цирроз печени, остеопороз, опухоли. При этих мутациях происходит сборка дисфункционального теломеразного комплекса, который сохраняет частичную активность, или фермента с полностью утраченной каталитической активностью. Обычно только одна аллель теломеразного гена отвечает за неактивность теломеразно-ферментативного комплекса.
Инактивация теломеразы приводит к мутациям в генах. Клинические проявления при недостаточности теломеразы могут быть различной природы. Недостаточность теломеразы не является причиной патологии, но отражается на поддержании гомеостаза теломер.
Ограничения теломеразной теории старения В настоящее время проводятся исследования связи между длиной теломер и старением у человека [16]. Однако уже сейчас по результатам этих работ можно сделать заключение о том, что длина теломер и скорость их укорочения сильно варьируют в популяции. Предполагается, что длина теломер выступает в качестве надежного биомаркера старения не на всех, а только на определенных стадиях жизни [17]. Остается открытым вопрос, является ли она показателем нормального процесса старения или только маркером заболеваний, связанных с ним [18]. Из-за недостаточности знаний о процессе старения до сих пор трудно это установить [19, 20]. Кроме того, пока отсутствуют научные работы о вероятном влиянии предшествующих заболеваний, таких как вирусные инфекции, на длину теломер, а также информация о том, нужно ли это рассматривать в качестве потенциальных факторов, которые будут влиять на оценку результатов. Возможно, длину теломер можно считать биомаркером старения только на определенных этапах жизни [21].
Связь длины теломер и некоторых заболеваний
Сердечно-сосудистые заболевания
У пациентов с более короткими лейкоцитарными теломерами наблюдалось повышение смертности от сердечно-сосудистых заболеваний в 3 раза, от инфекционных – в 8,5 раза [15].
В исследовании S. Brouilette и соавт. [22] было показано, что у молодых людей с более короткими теломерами риск развития преждевременного инфаркта миокарда оказался увеличен в 2,8-3,2 раза [22]. Зависимость сердечно-сосудистого риска от длины теломер наблюдали у пожилых людей [23]. Длина теломер лейкоцитов периферической крови имеет прогностическое значение при развитии сердечно-сосудистых заболеваний [24].
Хронический стресс
Для людей, живущих в условиях хронического стресса, характерна меньшая продолжительность жизни и раннее развитие возрастных болезней. Причиной стремительного укорочения теломер в данном случае может быть активация вегетативной нервной и нейроэндокринной систем с последующим выбросом глюкокортикоидных гормонов.
Наследственные дегенеративные заболевания
Результаты изучения особенностей хромосом пациентов с наследственными дегенеративными заболеваниями, характеризующимися преждевременным старением организма, в т. ч. аутосомно-доминантным врожденным дискератозом (синдром Цинссера-Энгмена-Коула), прогерией взрослых (синдром Вернера) и атаксией-телеангиэктазией (синдром Луи-Бар), указывают на важную роль, принадлежащую теломерам в процессах старения.
Приобретенные дегенеративные заболевания
Существуют доказательства роли теломер в развитии приобретенных дегенеративных заболеваний. Наиболее показательным примером является цирроз печени, при котором значительное повышение скорости обновления гепатоцитов сопровождается прогрессивным укорочением теломер. В результате клетки утрачивают способность к делению и погибают, что приводит к развитию печеночной недостаточности.
Риск развития слабоумия и сахарный диабет
В работах L. Hong и M. Sampson продемонстрирована взаимосвязь между длиной теломер и риском развития слабоумия, а также сахарным диабетом [25, 26].
Выводы
1. Концы линейных хромосом с 3`-конца ДНК заканчиваются повторяющимися последовательностями нуклеотидов – теломерами, которые синтезируются рибонуклеиновым ферментом теломеразой.
2. Структуры теломер одинаковы у всех позвоночных – (TTAGGG)n.
3. Укорочение теломер – ключевой фактор, запускающий развитие дегенеративных заболеваний и уменьшающий продолжительность жизни.
4. Длина теломер является прогностическим показателем риска заболеваний, их прогрессирования и преждевременной смертности, а также характеристикой снижения выживаемости у пациентов с ишемической болезнью сердца и инфекционными заболеваниями.
5. Измерение активности теломеразы может обеспечить более ранний прогноз геномной стабильности долгосрочной жизнеспособности, чем длина теломер.
6. Оценка различных аспектов состояния теломер может помочь в прогнозировании течения различных заболеваний и предоставить новые возможности для профилактических и терапевтических вмешательств, в т. ч. временной активации эндогенной теломеразы.
Список литературы находится в редакции.
Медична газета «Здоров’я України 21 сторіччя» № 5 (426), березень 2018 р.