


3 березня, 2019
Молекулярно-генетическая диагностика синдрома Жильбера
Впервые этот синдром был описан французским врачом A. Gilbert в начале XX века [1]. Частота встречаемости СЖ в европеоидной, негроидной и монголоидной расах составляет 5-10, 30-35 и 2-3% соответственно [2, 3]. По статистическим данным, указанная патология чаще проявляется у мужчин (12,4%), чем у женщин (4,8%; соотношение 3:1-4:1). Средняя концентрация билирубина при СЖ значительно выше у мужчин, что может быть связано с ингибированием андрогенными стероидами процесса ферментативного глюкуронирования билирубина. СЖ передается по аутосомно-рецессивному типу наследования и, как правило, манифестация наблюдается в возрасте до 30 лет. Основным клиническим проявлением СЖ является желтушное окрашивание кожи, склер, слизистых. Во время эпизодов желтухи отмечаются боль в животе, спазмы в желудке, вздутие живота, диарея или запор, чувство усталости, потеря аппетита, головокружение, депрессия, потливость. У пациентов возможна манифестация с развитием токсических реакций при приеме некоторых лекарственных препаратов: анаболических стероидов, глюкокортикоидов, андрогенов, рифампицина, циметидина, хлорамфеникола, стрептомицина, салицилата натрия, ампициллина, кофеина, этинилэстрадиола, парацетамола, иринотекана.
Источники билирубина
У здорового человека в сутки распадается около 1% циркулирующих эритроцитов. Освободившийся при этом гемоглобин в клетках ретикулоэндотелиальной системы (костный мозг, селезенка) распадается на гем (небелковую железосодержащую часть, в молекуле которой железо связано с протопорфирином) и белковое соединение – глобин. Основная часть билирубина (80-85%) синтезируется в печени, а около 15-20% поступает из других источников, таких как гемопротеины (миоглобин, цитохромы, каталаза, пероксидаза) и незрелые клетки (эритробласты, незрелые ретикулоциты). В организме человека образуется 250-350 мг билирубина ежедневно.
Ферментативный механизм образования билирубина
После распада гемоглобина в ретикулоэндотелиальной системе, образования гема и глобина происходит окисление гема под действием гемоксигеназы. В этой реакции расходуется три молекулы кислорода и никотинадениндинуклеотидфосфат с образованием эквивалентного количества биливердина и оксида углерода. В дальнейшем биливердин под действием биливердинредуктазы конвертируется в билирубин. Образовавшийся билирубин (неконъюгированный, непрямой, свободный) является гидрофобным, липофильным соединением, не способным к почечной и билиарной секреции. Даже умеренно повышенный уровень свободного билирубина является токсичным для центральной нервной системы, что проявляется невыраженными клиническими симптомами (ослабление внимания, утомляемость и др). Билирубин имеет высокое сродство к альбумину, поэтому 99% неконъюгированного билирубина циркулирует в плазме в виде комплекса с этими белками. Неконъюгированный билирубин, связанный с альбумином, транспортируется в печень, где происходит диссоциация этого комплекса. В гепатоцитах непрямой билирубин отщепляется от альбумина и взаимодействует с глюкуроновой кислотой при участии фермента уридиндифосфатглюкуронозилтрансферазы (UGT1A1), образуя при этом глюкуронид билирубина (моноглюкуронид – 15% и диглюкуронид – 85%) или конъюгированный (прямой) билирубин (рис. 1).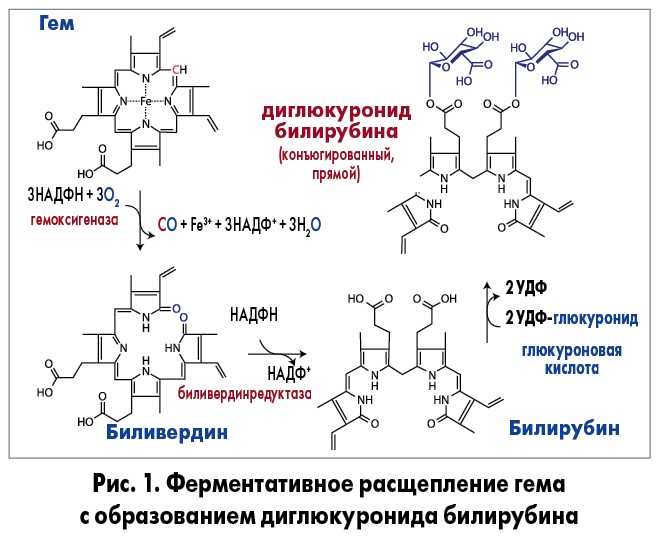
У большинства пациентов с СЖ гипербилирубинемия проявляется в молодом возрасте. В зависимости от референтных интервалов, установленных в лабораториях, верхний предел для общего билирубина в норме – от 17 до 20 мкмоль/л, из которых 75% приходится на неконъюгированный билирубин [4]. При СЖ концентрация общего билирубина составляет от 20 до 50 мкмоль/л и редко превышает 85 мкмоль/л.
Несмотря на развитие новых медицинских технологий, диагностика СЖ до сих пор остается достаточно сложной. При наличии огромного количества лабораторных (неконъюгированный билирубин, общий анализ крови с ретикулоцитами, ферменты печени) и функциональных (гипокалорийный, с никотиновой кислотой, с фенобарбиталом) тестов не выработан оптимальный алгоритм применения, основанный на их чувствительности и специфичности. В связи с этим рекомендуется проведение генетического анализа для подтверждения СЖ.
Клиническая картина СЖ обусловлена снижением до 30% уровня функциональной активности UGT1A1 – фермента, необходимого для конъюгирования билирубина. Этот фермент кодируется геном UGT1A, расположенным на 2q37 хромосоме.
Ген UGT1A
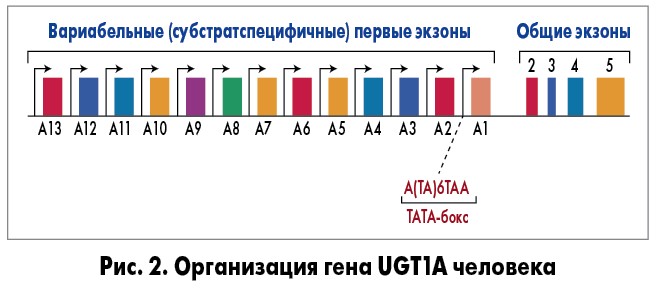 В 1991 г. J. Ritter и соавт. сообщили о выделении из печени двух клонов, кодирующих ДНК – UGT1A и UGT1D (переименованные в дальнейшем в UGT1A и UGT1A4), с билирубиновой глюкуронилтрансферазной активностью [5]. Нуклеотидная последовательность из 30 регионов этих двух клонов была одинаковой (кодируются одним геном), что и подтвердилось спустя год, когда авторы сообщили о выделении большого участка гена UGT1A с его уникальной структурой [6]. В начале 2000-х гг. был полностью изучен принцип работы гена UGT1A: имеет длину 218 kb и содержит 13 аминотерминальных экзонов, которые кодируют субстрат, связывающий домен и 4 общих карбокситерминальных экзонов, кодирующих УДФ-глюкуроновую кислоту [7].
В 1991 г. J. Ritter и соавт. сообщили о выделении из печени двух клонов, кодирующих ДНК – UGT1A и UGT1D (переименованные в дальнейшем в UGT1A и UGT1A4), с билирубиновой глюкуронилтрансферазной активностью [5]. Нуклеотидная последовательность из 30 регионов этих двух клонов была одинаковой (кодируются одним геном), что и подтвердилось спустя год, когда авторы сообщили о выделении большого участка гена UGT1A с его уникальной структурой [6]. В начале 2000-х гг. был полностью изучен принцип работы гена UGT1A: имеет длину 218 kb и содержит 13 аминотерминальных экзонов, которые кодируют субстрат, связывающий домен и 4 общих карбокситерминальных экзонов, кодирующих УДФ-глюкуроновую кислоту [7].
Вариабельные экзоны 1A2-1A13 не участвуют в метаболизме билирубина. Генетические мутации, связанные с отсутствием или снижением ферментативной активности, которые вызывают нарушения конъюгирования билирубина, локализованы в вариабельных экзонах 1А1 и общих экзонах с 2-го по 5-й (рис. 2).
UGT1A1
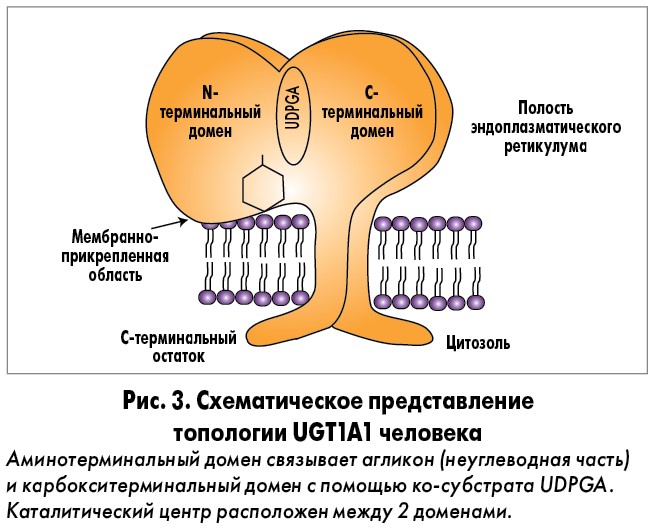 Ген UGT1A1 кодирует фермент UGT1A1. В печени последний катализирует реакцию соединения (конъюгирования) билирубина (непрямой, неконъюгированный) с глюкуроновой кислотой. СЖ обусловлен мутацией в гене UGT1A. Активность UGT1A1 у пациентов с этим синдромом составляет около 30% от нормы, основанной на исследованиях с использованием образцов печени человека [8, 9]. В настоящее время семейство ферментов UGT1A1 продолжают активно изучать. Изоформы UGT1A обнаружены в различных отделах организма человека: в печени – UGT 1А1, 1А3, 1А4, 1А6, 1А9, 1А7, в пищеводе и желудке – UGT 1А8, в кишечнике – UGT1А10. Функцией ферментов семейства UGT 1А1 является конъюгация как эндогенных метаболитов, гормонов, нейротрансмиттеров, так и экзогенных веществ (различных ксенобиотиков, канцерогенов, лекарственных препаратов) [10, 11]. Белки семейства UGT человека (ЕС 2.4.1.17) имеют средний молекулярный вес приблизительно 56 кДа; в их состав входит около 530 аминокислот. UGT являются интегральными мембранными белками эндоплазматического ретикулума, состоящими из 2 аналогичных по размеру доменов (рис. 3).
Ген UGT1A1 кодирует фермент UGT1A1. В печени последний катализирует реакцию соединения (конъюгирования) билирубина (непрямой, неконъюгированный) с глюкуроновой кислотой. СЖ обусловлен мутацией в гене UGT1A. Активность UGT1A1 у пациентов с этим синдромом составляет около 30% от нормы, основанной на исследованиях с использованием образцов печени человека [8, 9]. В настоящее время семейство ферментов UGT1A1 продолжают активно изучать. Изоформы UGT1A обнаружены в различных отделах организма человека: в печени – UGT 1А1, 1А3, 1А4, 1А6, 1А9, 1А7, в пищеводе и желудке – UGT 1А8, в кишечнике – UGT1А10. Функцией ферментов семейства UGT 1А1 является конъюгация как эндогенных метаболитов, гормонов, нейротрансмиттеров, так и экзогенных веществ (различных ксенобиотиков, канцерогенов, лекарственных препаратов) [10, 11]. Белки семейства UGT человека (ЕС 2.4.1.17) имеют средний молекулярный вес приблизительно 56 кДа; в их состав входит около 530 аминокислот. UGT являются интегральными мембранными белками эндоплазматического ретикулума, состоящими из 2 аналогичных по размеру доменов (рис. 3).
Полиморфизм UGT1A1
У человека подсемейства UGT1 и UGT2 содержат 19 различных изоферментов, имеющих близкую последовательность аминокислот [12, 13]. Однако только UGT1A1 способна глюкуронировать билирубин [14]. В 1995 г. было показано, что мутация в промоторном участке гена UGT1A1 является необходимой, но недостаточной для полной манифестации СЖ. Эта мутация, снижающая активность фермента UGT1A1, представляет собой вставку двух нуклеотидов в ТАТА-боксе промотора гена. В этом случае нормальная последовательность A(TA)6TAA трансформируется в последовательность A(TA)7TAA. Шесть повторов тимин-аденин (TA) в области промотора A(TA)6TAA соответствуют нормальной функциональной активности фермента UGT. При вставке седьмого ТА-повтора A(TA)7TAA ухудшается связывание с транскрипционным фактором, в результате чего уменьшается экспрессия гена, что ведет к снижению функциональной активности фермента, которая проявляется непрямой (неконъюгированной) гипербилирубинемией. При наличии вставки TA в гомозиготном состоянии наблюдается снижение активности фермента на 30% и конъюгации билирубина в гепатоцитах на 80% по отношению к норме. Тенденция к прогрессирующему снижению активности фермента также отмечается при увеличении TA-повторов (например, до 8). В случае возрастания числа ТА-повторов до 8 в одном из аллельных генов или в обоих СЖ также считается подтвержденным – А(ТА)7 ТАА / А(ТА)8ТАА и А(ТА)8 ТАА / А(ТА)8ТАА. Активность фермента UGT1A1 находится в обратной зависимости от количества повторов.
К настоящему времени для UGT1A1 показано существование не менее 4 аллелей: A(TA)6TAA (рис. 4).
- UGT1A1*1 – «дикий тип», wild type;
- UGT1A1*28 – аллель A(TA)7TAA, наиболее частый мутантный;
- UGT1A1*36 – аллель (TA)5;
- UGT1A1*37 – аллель (TA)8.
Частота мутантного аллеля UGT1A1*28 в различных популяциях следующая: 26-39% – для европеоидной, 9-33% – для азиатской, 40-56% – для негроидной популяции. Другие мутантные аллели – UGT1A1*36 и UGT1A1*37 – обнаружены главным образом в негроидной популяции с частотой 2-16%.
Идентификация количества ТА-повторов в промоторной области гена UGT1A1
Самым быстрым и эффективным методом для выявления СЖ является ДНК-диагностика с использованием метода полимеразной цепной реакции (ПЦР), который заключается в определении числа TA-повторов в гене UGT1A1.
Метод ПЦР с использованием капиллярного гель-электрофореза
 Молекулярно-генетическая диагностика проводится методом ПЦР с меченными карбоксифлуоресцеином праймерами с разделением продуктов амплификации при помощи капиллярного гель-электрофореза.
Молекулярно-генетическая диагностика проводится методом ПЦР с меченными карбоксифлуоресцеином праймерами с разделением продуктов амплификации при помощи капиллярного гель-электрофореза.
Ампликон длиной 96 пар основ соответствует участку промотора гена UGT1A1 с 7 ТА-повторами, что позволяет подтвердить диагноз СЖ. Ампликон длиной 94 пары основ соответствует нормальному участку промотора гена UGT1A1 с 6 ТА-повторами. Наличие двух ампликонов длиной 96 и 94 пары основ свидетельствует о гетерозиготном носительстве 6/7 ТА-повторов.
Секвенирование продуктов ПЦР
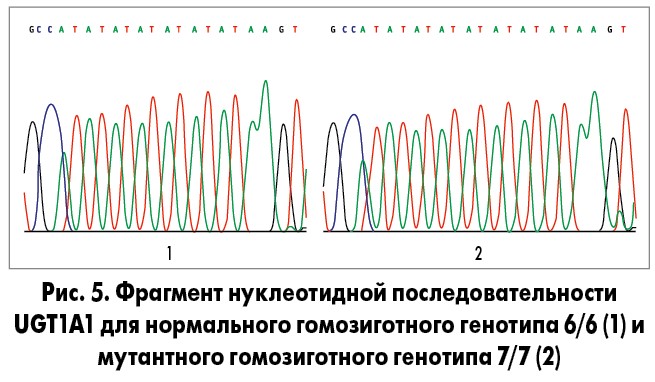
На рисунке 5 представлен фрагмент нуклеотидной последовательности UGT1A1 для нормального гомозиготного генотипа 6/6 (структура повтора: A-(TA)6-TAA, результат секвенирования продукта ПЦР размером 97 пар нуклеотидов) и фрагмент нуклеотидной последовательности UGT1A1 для мутантного гомозиготного генотипа 7/7 (структура повтора: A-(TA)7-TAA, результат секвенирования продукта ПЦР размером 99 пар нуклеотидов).
Интерпретация результатов исследования
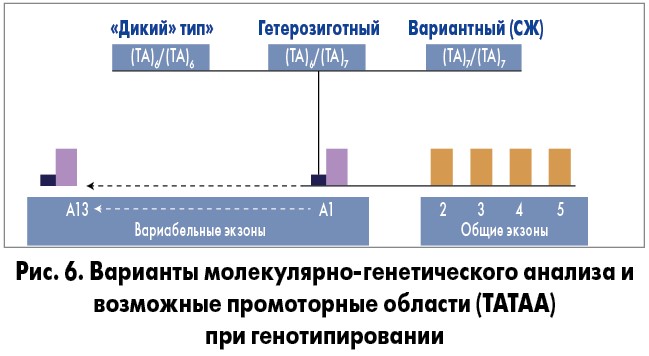
Варианты молекулярно-генетического анализа могу быть следующими (рис. 6):
- UGT1A1 (ТА)6/(ТА)6 – нормальный («дикий») генотип. CЖ не выявлен.
- UGT1A1 (ТА)6/(ТА)7 – генотип с увеличением ТА-повторов в гене в гетерозиготном состоянии. Риск развития СЖ – в латентной или легкой форме.
- UGT1A1 (ТА)7/(ТА)7 – генотип с увеличением ТА-повторов в гене в гомозиготном состоянии. Очень большой риск развития СЖ. Заболевание может протекать со значительным повышением уровня билирубина и тяжелыми клиническими проявлениями.
Выводы
 В настоящее время все большее значение приобретает необходимость лабораторного подтверждения СЖ, что связано с установлением дифференциального диагноза. При выявлении повышенного уровня билирубина в сыворотке крови в течение продолжительного времени рекомендуется прежде всего выполнить молекулярно-генетический анализ. Назначение исследования ТА-повторов в гене UGT1A важно в следующих случаях: при дифференциальной диагностике СЖ и других заболеваний, сопровождающихся гипербилирубинемией, перед началом лечения с использованием лекарственных препаратов, обладающих гепатотоксическим действием, при возможности осложнений терапии иринотеканом, слабовыраженной неинфекционной желтухе, отягощенном семейном анамнезе и др. Необходимость назначения молекулярно-генетического анализа и интерпретации полученных результатов определяется врачом-генетиком при проведении медико-генетического консультирования.
В настоящее время все большее значение приобретает необходимость лабораторного подтверждения СЖ, что связано с установлением дифференциального диагноза. При выявлении повышенного уровня билирубина в сыворотке крови в течение продолжительного времени рекомендуется прежде всего выполнить молекулярно-генетический анализ. Назначение исследования ТА-повторов в гене UGT1A важно в следующих случаях: при дифференциальной диагностике СЖ и других заболеваний, сопровождающихся гипербилирубинемией, перед началом лечения с использованием лекарственных препаратов, обладающих гепатотоксическим действием, при возможности осложнений терапии иринотеканом, слабовыраженной неинфекционной желтухе, отягощенном семейном анамнезе и др. Необходимость назначения молекулярно-генетического анализа и интерпретации полученных результатов определяется врачом-генетиком при проведении медико-генетического консультирования.
В Украине анализ для определения ТА-повторов в промоторной области гена UGT1A1, ответственного за развитие СЖ, возможно сделать в некоторых частных медицинских лабораториях.
Список литературы находится в редакции.
Медична газета «Здоров’я України 21 сторіччя» № 24 (445), грудень 2018 р.