


12 липня, 2016
Метформин: новые горизонты применения (II часть)
Продолжение. Начало в № 10.
Метформин и рак
AMPK-зависимые механизмы
Противоопухолевые эффекты метформина в значительной степени опосредуются активацией сигнального пути LKB1/AMPK. LKB1 является опухолевым супрессором, и мутация гена LKB1 ассоциируется с синдромом Пейта-Йегерса – врожденным заболеванием с предрасположенностью к злокачественным новообразованиям. Было продемонстрировано, что протекторный эффект метформина ослабляется при фармакологическом ингибировании AMPK и в экспериментальных моделях у AMPK-нокаутированных животных. Активация AMPK приводит к подавлению передачи сигналов в пути mTOR (мишени рапамицина млекопитающих) – нутриентчувствительного регулятора синтеза белка, роста и пролиферации клеток. Этот эффект достигается путем прямого фосфорилирования Ser1345 и Thr1227 опухолевого супрессора TSC2 (белка туберозного склероза 2), который формирует mTOR-комплекс 1 (mTORC1) с TSC1. Кроме того, AMPK может предотвращать активацию mTORC1 посредством фосфорилирования его связующего партнера Raptor.
Активация AMPK сопровождается подавлением эффекта Варбурга – приобретаемого раковыми клетками метаболического фенотипа, который характеризуется предпочтением аэробного гликолиза над окислительным фосфорилированием. Эта трансформация облегчается гипоксия-индуцибельным фактором 1α – транскрипционным фактором, активирующим экспрессию гликолитического гена в ответ на передачу сигналов mTORC1. Классической метаболической характеристикой рака, связанной с эффектом Варбурга, является повышенный синтез жирных кислот de novo и повышенные уровни ключевого липогенного фермента – синтетазы жирных кислот (FAS), описанные при различных типах рака у человека. Установлено, что активация AMPK снижает экспрессию FAS в клетках рака предстательной железы и уменьшает их выживаемость. В экспериментальном исследовании на модели карциномы толстой кишки было продемонстрировано, что метформин снижает экспрессию FAS и противодействует стимулирующему влиянию высококалорийной пищи на рост опухоли. Эти данные свидетельствуют о том, что метформин может проявлять свои противоопухолевые эффекты путем изменения метаболизма глюкозы и липидов, что в конечном итоге ограничивает доступ раковых клеток к субстратам, необходимым для роста.
Недавно в нескольких работах было установлено, что метформин может действовать как антифолатный препарат, нарушая фолатный цикл подобно аналогичному эффекту антифолатных химиотерапевтических агентов. Фолатный, или одноуглеродный, цикл является ключевым регулятором клеточного метаболизма и интегратором нутриентного статуса. Он ассимилирует глюкозу и аминокислоты, которые посредством химических реакций трансформируются для выполнения различных биологических функцией. Последние включают клеточный биосинтез, регуляцию кислотно-основного состояния, регуляцию эпигенетики путем метилирования нуклеиновых кислот и белков, сохранение генома посредством регуляции пула нуклеотидов. Например, фолатный цикл регулирует de novo синтез нуклеотидов, необходимых для починки и репликации ДНК, а также для продукции универсального донатора метильной группы S-аденозилметионина, принимающего участие в различных реакциях метилирования, в частности метилирования ДНК. Химиотерапевтические препараты, такие как метотрексат, аминопретин и азасерин, блокируют фолатный метаболизм и нарушают de novo синтез тимидилатных и пуриновых нуклеотидов. Установлено, что метформин нарушает метаболизм фолата и метионина у Escherichia coli, а также в клетках рака грудной железы с одновременным снижением метаболитов глутатиона и триптофана.
Помимо вышеописанных изменений метаболизма метформин может регулировать клеточный цикл путем взаимодействия с классическими онкогенами и опухолевыми супрессорами. Например, было продемонстрировано, что метформин вызывает AMPK-зависимую понижающую регуляцию c-MYC в клеточных линиях рака грудной железы. Этот эффект опосредуется повышенной экспрессией miR‑33a (микроРНК, которая связывается с регионом 3’UTR с-MYC), что, в свою очередь, вызывается повышающей регуляцией фермента РНКазы III Dicer в ответ на лечение метформином. Модуляция Dicer под действием метформина имеет особую значимость с учетом того, что низкие уровни этого фермента ассоциируются с неблагоприятным прогнозом при раке яичников, грудной железы и легких.
AMPK-независимые механизмы
Описан также ряд AMPK-независимых механизмов, объясняющих противоопухолевые свойства метформина. В частности, метформин может защищать ДНК от повреждений и мутаций, подавляя продукцию АФК комплексом I. Кроме того, может активировать mTORC1 в отсутствие AMPK, что достигается ингибированием Rag-ГТФаз, отвечающих за индуцирование транслокации mTORC1 в клеточные компартменты, содержащие активатор mTORC1 – Rheb (гомолог Ras, концентрирующийся в головном мозге). Антипролиферативные эффекты метформина при раке предстательной железы связаны с AMPK-независимым ингибированием критического регулятора клеточного цикла циклина D1. Предполагается, что данный процесс опосредуется р53-зависимой повышающей регуляцией REDD1 (белка, регулируемого при эмбриогенезе и повреждении ДНК). Исследование, проведенное на клетках плоскоклеточного рака пищевода, показало, что метформин способствует их апоптозу и аутофагии, эффективно подавляя рост опухоли. Этот эффект был обусловлен инактивацией сигнального пути Stat3 (сигнальный трансдуктор и активатор транскрипции 3) / Bcl2. На другой клеточной модели были продемонстрировано, что метформин нарушает захват глюкозы клетками рака легкого путем аллостерического ингибирования гексокиназы-II. Дефицит этого энергетического субстрата вызывает деполяризацию митохондрий с последующим апоптозом. Аналогичный эффект также наблюдался в отношении клеток рака грудной железы.
Комбинированная противоопухолевая терапия с включением метформина
Получены доказательства, что метформин улучшает ответ раковых клеток на лучевую терапию, а также повышает чувствительность различных типов раковых клеток к широко применяющимся химиотерапевтическим средствам, включая цисплатин, паклитаксел, карбоплатин и доксорубицин. Однако следует отметить, что метформин снижал цитотоксический эффект цисплатина на клеточных линиях глиомы, нейробластомы, фибросаркомы и лейкоза посредством AMPK-независимой регуляции сигнального пути выживания Akt. Метформин может быть особенно эффективным при использовании в комбинации с низкомолекулярными ингибиторами киназы. На клеточных линиях меланомы было продемонстрировано, что эффективность ингибиторов протеинкиназы BRAF (гомолога В1 вирусного онкогена мышиной саркомы v Raf) ограничивается компенсаторным окислительным фосфорилированием. Соответственно, ингибиторы окислительного фосфорилирования повышают эффективность этих препаратов. В отличие от многих ингибиторов окислительного фосфорилирования, проявляющих выраженную токсичность, метформин имеет благоприятный профиль безопасности и, следовательно, высокий терапевтический потенциал.
Метформин и старение
Биологическая основа процесса старения остается главной загадкой для науки, однако за последние годы на пути к ее решению был достигнут значительный прогресс. Генетические, диетические и фармакологические вмешательства могут улучшать здоровье и продлевать жизнь ряда лабораторных организмов, таких как нематода Caenorhabditis elegans, плодовая муха Drosophila melanogaster и домовая мышь Mus musculus. Эти вмешательства ослабляют процесс старения и увеличивают продолжительность жизни, защищая от возрастных патологических изменений. Например, контролированное снижение потребления пищи без нутритивной недостаточности продлевает жизнь многим животным. Метформин может имитировать этот эффект и таким образом обеспечивать схожие преимущества у человека.
Бигуаниды и выживаемость C. elegans
В многочисленных исследованиях было установлено, что бигуаниды могут поддерживать и продлевать жизнь C. elegans. В одной из первых таких работ назначение буформина в дозе 0,1 мг/мл на протяжении жизни (включая личиночную стадию) увеличивало среднюю и максимальную продолжительность жизни взрослого червя на 23 и 26% соответственно. Назначение метформина на протяжении жизни в дозе 50 ммоль увеличивало среднюю продолжительность жизни на 40%. Интересно, что более низкие и более высокие дозы метформина (10 и 100 ммоль соответственно) не имели подобного эффекта. В двух недавних исследованиях было продемонстрировано, что эффекты метформина в дозе 50 ммоль не зависят от его назначения на ранних стадиях развития и увеличение выживаемости также достигается при использовании препарата в более поздние периоды жизни. Аналогичным образом фенформин в концентрациях 1,5; 3 и 4,5 ммоль продлевал жизнь взрослых червей на 5, 21 и 26% соответственно. Так как же бигуаниды увеличивают продолжительность жизни C. elegans?
Метформин обладает комплексным механизмом действия, вызывая широкий спектр физиологических эффектов. Установлено, что способность препарата продлевать жизнь в значительной степени связана с его влиянием на метаболизм фолата и метионина бактерий-симбионтов. Кроме того, метформин действует как прямой метаболический стрессор (вероятно, путем ингибирования комплекса I дыхательной цепи), активируя ответ на окислительный стресс и детоксикацию с вовлечением ряда метаболических медиаторов, включая AMPK и SKN‑1. Это подтверждается тем, что назначение антиоксидантов, таких как N-ацетилцистеин, препятствующих продукции АФК и мутациям AMPK/SKN‑1, нарушает развитие адекватного детоксицирующего ответа, индуцируемого метформином путем влияния на бактериальный метаболизм.
Метформин может не только продлевать жизнь, но и увеличивать продолжительность здоровой жизни C. elegans. Так, меторфмин снижает накопление пигмента молекулярного повреждения липофусцина, увеличивает подвижность на поздних периодах жизни, снижает накопление жировой ткани, ослабляет возрастные морфологические изменения и повышает выживаемость особей, подвергающихся длительной аноксии.
Бигуаниды и выживаемость насекомых
У насекомых эффекты бигуанидов на выживаемость имеют свои особенности. Назначение метформина в дозах 1; 2,5 и 5 ммоль не влияло на выживаемость мужских и женских особей D. melanogaster, при этом более высокие дозы (100 ммоль у самцов; 25, 50 и 100 ммоль у самок) снижали этот параметр, несмотря на выраженную активацию AMPK и снижение жировых запасов. Этот результат противоречит данным других работ, где было продемонстрировано, что повышенные уровни экспрессии и активации AMPK увеличивают выживаемость C. elegans и D. melanogaster.
Хорошо описанной особенностью предиабета у человека является инфекционно-индуцируемая гипергликемия. На модели ожирения у D. melanogaster назначение метформина в дозе 10 ммоль снижало неблагоприятные последствия инфекции Rhizopus – гриба, вызывающего мукормикоз у пациентов с диабетом. Метформин уменьшал набор веса, нормализовал уровни глюкозы, повышенные вследствие инфекции, и повышал выживаемость. При назначении в дозе 5 ммоль метформин замедлял возрастные изменения кишечных стволовых клеток D. melanogaster, что проявлялось в снижении молекулярных маркеров повреждения ДНК gH2AX foci и 8-oxodG. В одном из последних исследований, выполненных на домовых сверчках Acheta domesticus, метформин увеличивал среднюю и максимальную продолжительность жизни у 23,2% мужских и 43,7% женских особей.
Бигуаниды и выживаемость грызунов
Результаты многочисленных исследований, проведенных на грызунах, указывают на эволюционно сохранившуюся жизнепродлевающую роль бигуанидов. Назначение метформина мышам линий HER‑2/neu, SHR, 129/Sv, C57BL/6 и B6C3F1 в широком спектре дозировок увеличивало продолжительность жизни на 4-38%. При этом установлено, что данный эффект препарата зависит от генотипа, пола, используемого бигуанида и его дозы, а также от длительности лечения. В целом, чем позже было начато лечение, тем менее выраженным было продление жизни (схожая зависимость наблюдалась и в экспериментах с C. elegans). Например, назначение мышам SHR метформина в 15-месячном возрасте не увеличивало продолжительность жизни по сравнению с назначением в 3-месячном возрасте (0 vs 38% соответственно). У мышей C57BL/6 доза метформина 0,1% от суммарного рациона продлевала жизнь на 5,83%, тогда как доза 1% – сокращала ее на 14,4%; уменьшение продолжительности жизни было обусловлено нефротоксичностью. У мышей 129/Sv назначение метформина продлевало жизнь на 5% у самок, но не у самцов. Интересно, что влияние бигуанидов на продолжительность жизни крыс менее выражено по сравнению с мышами.
Замедление старения и продление жизни лабораторных животных при назначении метформина, по-видимому, обусловлено широким спектром эффектов препарата, включая изменения веса и температуры, нормализацию сывороточных уровней глюкозы, инсулина, триглицеридов и холестерина, регуляцию эстрогенной функции и антинеопластические эффекты. Недавно в исследовании Martin-Montalvo и соавт. было установлено, что метформин, имитируя эффекты ограничения энергетической ценности рациона, играет важную роль в продлении жизни и сохранении здоровья. Метформин улучшает фитнес, повышает чувствительность тканей к инсулину, снижает уровни липопротеинов низкой плотности и холестерина без снижения калорийности пищи. Интересно, что повышение активности AMPK при назначении метформина происходит без сопутствующих изменений в дыхательной цепи. Кроме того, метформин активирует антиоксидантные и противовоспалительные ответы, что уменьшает окислительное повреждение тканей и хроническое воспаление.
В целом накопленные экспериментальные данные сформировали базис для клинического применения метформина как антиэйджингового препарата. Продолжение исследований в этой области, очевидно, имеет огромные перспективы.
Метформин и увеличение продолжительности жизни человека
На сегодня проспективные исследования, в которых оценивалось бы влияние метформина на выживаемость здоровых людей, не проводились. В 10-летнем рандомизированном клиническом исследовании метформина у пациентов с избыточной массой тела или ожирением, страдающих СД 2 типа (UKPDS-O), были продемонстрированы долгосрочные благоприятные эффекты препарата на здоровье и выживаемость, при этом наблюдалось снижение кардиальной и общей смертности у пациентов, получавших метформин. Важно отметить, что метаболические преимущества от лечения метформином длительно сохраняются даже после его отмены. Тем не менее молекулярные механизмы, посредством которых метформин проявляет отдаленные эффекты на выживаемость, не связанные с гликемическим контролем, остаются неизученными. Интересно, что в исследовании UKPDS-S, продолжавшемся 6,6 года, комбинированная терапия меторфмином и производным сульфонилмочевины (ПСМ) по сравнению с монотерапией ПСМ в разнородной группе пациентов с СД 2 типа (как с нормальной массой тела, так и с избыточным весом и ожирением) проявляла потенциально негативные эффекты. Учитывая эти данные и результаты экспериментальных работ, для лучшего понимания долгосрочных благоприятных эффектов метформина на продолжительность жизни человека целесообразно проведение новых исследований, нацеленных на здоровую и относительно молодую популяцию.
Метформин и микробиота
Практически все живые организмы не живут в изоляции, а формируют тесные взаимоотношения с другими видами, прежде всего микроорганизмами. Кишечник является домом для большинства микробов, населяющих тело человека, и эта широкая популяция бактерий, которую называют «забытым органом», принимает участие во многих метаболических процессах с важными последствиями для физиологии и модуляции метаболических фенотипов человека. На сегодня доказана связь между изменениями кишечной микробиоты (вызванными диетой, антибиотикотерапией и т. п.) и развитием таких заболеваний, как ожирение, СД 2 типа, метаболический синдром и онкопатология, а также ускоренным старением, т. е. микробиота может служить биомаркером здоровья макроорганизма. Следовательно, препараты, широко применяющиеся в лечении метаболических заболеваний, ассоциированных с дисфункцией микробиоты, могут влиять на бактериальные популяции и таким образом улучшать здоровье человека (рис. 3). Вопрос о том, в какой степени изменения микробиоты объясняют благоприятные эффекты метформина при метаболических заболеваниях, остается открытым. Предполагается, что этот препарат может модулировать состав и функции бактерий, населяющих гастроинтестинальный тракт.
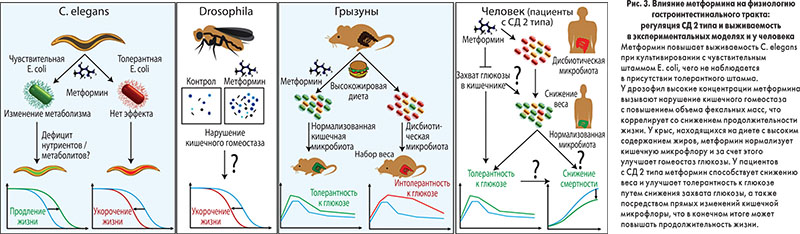
Метформин изменяет метаболизм и регулирует физиологию C. elegans
Нематода C. elegans сформировала выгодные взаимоотношения с E. coli, отражающие, хотя и в упрощенном виде, подобные ассоциации «здоровых» кишечных бактерий у человека. Было установлено, что у С. elegans метформин изменяет бактериальный метаболизм фолата и метионина, имитируя действие ограничения энергетической ценности пищи. При этом метформин замедляет процесс старения только в присутствии E. coli. Интересно, что у человека метформин также может вызывать дефицит фолата и витамина В12 с сопутствующим повышением уровней гомоцистеина.
Метформин влияет на кишечный гомеостаз у насекомых
Пищеварительный тракт D. melanogaster является одним из наиболее изученных органов этого насекомого. Как и у людей, задержка и абсорбция жидкости, перистальтика, кишечный транзит, частота дефекации и характер стула у дрозофил подвергаются сложной гомеостатической регуляции. В предыдущих исследованиях было установлено, что микробиом D. melanogaster модулирует развитие и метаболический гомеостаз организма-хозяина посредством влияния на сигнальный путь инсулина, тем не менее точная роль бактерий пока не определена.
Влияние метформина на функцию пищеварительного тракта у грызунов
Исследования на мышах показали, что метформин накапливается в стенке кишки в очень высоких концентрациях, и это может объяснять эффекты препарата на организм хозяина. В целом результаты исследований на грызунах указывают на то, что кишечник является главным органом, регулирующим действие метформина. Установлено, что метформин не только замедляет абсорбцию глюкозы в пищеварительном тракте, но и способствует ее утилизации путем неокислительного анаэробного метаболизма. Лактат как продукт анаэробного метаболизма по портальной вене может поступать в печень и там утилизироваться как субстрат для глюконеогенеза. Следовательно, кишечник играет важную роль в глюкозоснижающих эффектах метформина и одновременно служит защитным механизмом, предотвращающим гипогликемию.
Большинство микробов, сосуществующих с организмом-хозяином, находятся в пищеварительном тракте и поэтому подвергаются селективному воздействию высоких концентраций метформина. СД 2 типа и ожирение характеризуются структурными и функциональными изменениями популяций кишечных микробов, воспалительным состоянием и нарушением барьерной функции кишечника. Схожесть эффектов микробиоты и метформина на ожирение и регуляцию глюкозы крови указывает на существование общих механизмов, при этом противодиабетические эффекты метформина достигаются как прямым путем, так и опосредованно благодаря влиянию на кишечную микрофлору.
В двух недавно завершившихся исследованиях было установлено, что у мышей, находящихся на диете с высоким содержанием жиров, метформин улучшает метаболические маркеры и приближает состав фекальной микробиоты к таковому у животных, получающих нормальное питание. С помощью метагеномного анализа было продемонстрировано, что метформин вызывает глубокий сдвиг специфических подгрупп бактерий, в частности увеличивает содержание микроорганизмов рода Akkermansia. Кроме того, метформин повышает количество бокаловидных клеток, продуцирующих муцин, который является источником питания для Akkermansia municiphila. Известно, что назначение этой бактерии в качестве пробиотического препарата ассоциируется с улучшением метаболического профиля, снижением метаболической эндотоксемии и воспаления жировой ткани, улучшением гликемического контроля и снижением инсулинорезистентности; все эти эффекты также свойственны метформину.
У мышей, получающих обогащенную жирами пищу, метформин также увеличивает популяцию лактобацилл. Lactobacillus является одной из многих бактерий, которые могут утилизировать глюкозу с образованием лактата. Примечательно, что улучшение гомеостаза глюкозы при назначении метформина существенно ослабляется при сопутствующем применении антибиотиков широкого спектра действия. Эти данные свидетельствуют о том, что кишечная микробиота является терапевтической мишенью для метформина. По-видимому, повышенные уровни лактата и других субстратов для глюконеогенеза (пирувата, аланина) при лечении метформином не только защищают макроорганизм от гипогликемии, но и модифицируют структуру микробиоты, предоставляя новые энергетические субстраты определенным бактериям.
В исследовании на крысах было установлено, что комбинированная терапия пребиотическими пищевыми волокнами, такими как олигофруктоза, и метформином улучшает метаболический профиль у животных, находящихся на диете с высоким содержанием жиров. При этом олигофруктоза и метформин проявляли синергическое и аддитивное благоприятное действие на массу тела, печеночные триглицериды, секрецию глюкозозависимого инсулинотропного полипептида и лептина, а также на активацию AMPK, при этом значительно увеличивалось содержание бифидогенных бактерий, таких как Bifidobacterium spp.
Влияние метформина на кишечник и кишечную микрофлору человека
Терапевтические дозы большинства противодиабетических препаратов, использующихся у пациентов с СД 2 типа, находятся в диапазоне микрограмм и миллиграмм. В то же время эффективные дозы метформина могут достигать 2,5 г/сут. Тонкий кишечник является основным местом всасывания и аккумуляции метформина у животных и человека. Время абсорбции Т1/2 составляет 0,9-2,6 ч и зависит от принимаемой пищи, которая может отсрочивать этот процесс примерно на 35 мин. Метформин является стабильным и не связывается с белками, биодоступность составляет 50-60%. После перорального приема метформин достигает максимальной концентрации в периферической плазме через 2 ч, относительно быстро элиниминируется (период полувыведения – 1,7-4,5 ч) и экскретируется в неизмененном виде с калом и мочой. Как и у грызунов, метформин создает высокие концентрации в стенке тонкого кишечника человека, в 30-300 раз превышающие концентрации в плазме и других тканях. Концентрация метформина в просвете кишки после приема дозы 850 мг у пациентов с диабетом составляет около 20 ммоль. В целом эти данные свидетельствуют о том, что эффекты метформина на тонкий кишечник могут отличаться от таковых на другие ткани и также могут объяснять профиль побочных эффектов, свойственных этому препарату.
Наиболее распространенным побочным эффектом, ассоциированным с назначением метформина, является диспепсия. Лактатацидоз при лечении метформином регистрируется очень редко (3-9 на 100 тыс. пациенто-лет), и еще реже он вызывает фатальный исход (2-4 на 100 тыс. пациенто-лет). С другой стороны, диспепсические симптомы появляются на раннем этапе терапии, уменьшаются по мере продолжения лечения и полностью разрешаются после его отмены. Эти симптомы, как правило, имеют легкий характер и могут включать диарею, боль в животе, тошноту и рвоту, дисгевзию, вздутие и запор. Точные механизмы их развития не установлены. Предполагается, что они могут возникать вследствие изменений метаболизма серотонина, инкретинов, глюкозы и желчных кислот.
В недавних исследованиях также было продемонстрировано, что метформин изменяет микробный метаболизм в пищеварительном тракте. Это новая интересная гипотеза, которая может объяснять индивидуальные различия в эффектах препарата при диспепсических расстройствах. Karlsson и соавт. изучали состав и функцию фекальной микробиоты у 145 получавших метформин 70-летних европейских женщин с нормальным обменом глюкозы, нарушенной толерантностью к глюкозе и диабетом. Метагеномный анализ пациенток с СД 2 типа показал повышенные уровни представителей семейства Enterobactariaceae (Escherichia, Shigella, Klebsiella и Salmonella) и сниженные уровни Clostridium и Eubacterium. При этом наблюдалась статистически значимая корреляция между cодержанием E. coli и уровнем глюкагоноподобного пептида. Секреция последнего, как известно, снижена у пациенток с СД 2 типа, тогда как метформин повышает его уровни в плазме.
Другими побочными эффектами, ассоциированными с длительной терапией метформином, которые могут объясняться его влиянием на кишечную микробиоту, являются дефицит витамина В12 и фолата, а также изменение метаболизма желчных кислот. Бактерии, в частности Bifidobacterium, являются важными источниками витаминов для макроорганизма. Кроме того, многие анаэробные бактерии участвуют в трансформации желчных кислот в тонком кишечнике, главная функция которых состоит в облегчении метаболизма пищевых жиров и абсорбции жирорастворимых витаминов. В исследовании Napoliatano и соавт. отмена метформина у пациентов с СД 2 типа, находившихся на монотерапии этим препаратом, сопровождалась снижением уровней активного и общего глюкагоноподобного пептида‑1 и повышением сывороточных уровней желчных кислот, особенно холевой кислоты и ее конъюгатов. При возобновлении терапии метформином эти эффекты нивелировались. Кроме того, изменения желчных кислот положительно коррелировали с содержанием Firmicutes и отрицательно – с уровнем Bacteroidetes. В целом результаты этого и других современных исследований подтверждают данные ранних работ, в которых внутривенное назначение метформина, несмотря на достижения терапевтически эффективных концентраций в крови, не улучшало контроль глюкозы в отличие от перорального приема препарата.
Выводы
Несмотря на то что метформин на протяжении более полувека широко применяется как эффективный и безопасный препарат для лечения СД 2 типа, открываются все новые и новые механизмы его действия. Антигликемические свойства метформина опосредуются преимущественно подавлением глюконеогенеза в печени, и на сегодня считается, что данный эффект достигается путем ингибирования комплекса I в дыхательной цепи митохондрий. Последующее снижение энергетического статуса клетки непосредственно уменьшает глюконеогенез, снижает действие глюкагона и способствует активации AMPK – главного регулятора клеточного метаболизма; тем не менее пока не установлено, какой из этих трех эффектов вносит наибольший вклад в терапевтическое действие метформина. Кроме того, появляется все больше данных, свидетельствующих о том, что комплекс I является не единственной мишенью метформина; например, недавно было доказано, что метформин ингибирует митохондриальную глицерофосфатдегидрогеназу. Существует высокая вероятность того, что в ближайшем будущем будут идентифицированы новые мишени метформина. Исследование по С. elegans подчеркивает необходимость учитывать действие препаратов не только на макроорганизм, но и на его микробиоту. Бактериальные мишени метформина остаются малоизученными; по-видимому, микробиота посредством неких механизмов может регулировать ряд эффектов препарата на физиологию человека.
В настоящее время метформин одобрен только для лечения СД 2 типа, однако результаты многочисленных исследований, проведенных за последние годы, указывают на огромный терапевтический потенциал этого препарата при других заболеваниях. В настоящем обзоре приводятся различные механизмы, которые могут объяснять эти благоприятные эффекты; в то же время много вопросов остается нерешенными. Очевидно, что лучшее понимание молекулярных путей, задействованных в реализации эффектов метформина, расширит терапевтические горизонты применения этого препарата.
Список литературы находится в редакции.
Pryor R., Cabreiro F. Repurposing metformin: an old drug with new tricks in its binding pockets.Biochem J. 2015 Nov 1; 471 (3): 307-22.
Перевел с англ. Алексей Терещенко