


27 січня, 2026
Криза антибіотикорезистентності в Україні: актуальні терапевтичні рішення для сучасних викликів
За матеріалами конференції
 Незважаючи на те що Всесвітня організація охорони здоров’я визначає антибіотикорезистентність, або антимікробну резистентність (АМР), як одну із ключових глобальних загроз для людства, із позицій біології цей феномен є цілком логічною та еволюційно обґрунтованою відповіддю бактерій на тривалий і масований вплив ксенобіотиків, у тому числі й антибіотиків. Саме цій тематиці була присвячена доповідь доктора біологічних наук, провідного наукового співробітника Інституту молекулярної біології і генетики НАН України Олени Володимирівни Мошинець, представлена 28 листопада в межах 27-го засідання міждисциплінарного загальноакадемічного семінару в галузі природничих наук «Актуальні питання фізико-хімічної та математичної біології».
Незважаючи на те що Всесвітня організація охорони здоров’я визначає антибіотикорезистентність, або антимікробну резистентність (АМР), як одну із ключових глобальних загроз для людства, із позицій біології цей феномен є цілком логічною та еволюційно обґрунтованою відповіддю бактерій на тривалий і масований вплив ксенобіотиків, у тому числі й антибіотиків. Саме цій тематиці була присвячена доповідь доктора біологічних наук, провідного наукового співробітника Інституту молекулярної біології і генетики НАН України Олени Володимирівни Мошинець, представлена 28 листопада в межах 27-го засідання міждисциплінарного загальноакадемічного семінару в галузі природничих наук «Актуальні питання фізико-хімічної та математичної біології».
Ключові слова: антибіотикорезистентність, антимікробна резистентність, карбапенеми, макроліди, біоплівка.
Антибіотикорезистентність як біологічний феномен: взаємодія фенотипу та генотипу
Поява АМР зазвичай слідує за впровадженням відповідного класу антибіотиків, однак не є наслідком виключно некоректного або надмірного застосування останніх, а відображає адаптаційні можливості бактерій.
Так, гени стійкості до антибіотиків існували в бактеріальних популяціях задовго до початку ери антибіотикотерапії. Зокрема, ферменти, що інактивують β-лактамні антибіотики, еволюційно сформувалися з білків, які первинно виконували інші біологічні функції, зокрема функції протидії хімічному «спілкуванню» конкурентів в екологічній ніші, а генетичні детермінанти стійкості до аміноглікозидів в ентерококів були присутні ще до появи цих препаратів у клінічній практиці.
АМР є багатокомпонентним біологічним феноменом, який не може бути зведений до єдиного механізму. Вона формується на перетині трьох взаємопов’язаних складових, а саме фенотипової, вродженої генетичної й набутої генетичної резистентності (рис. 1) [1].
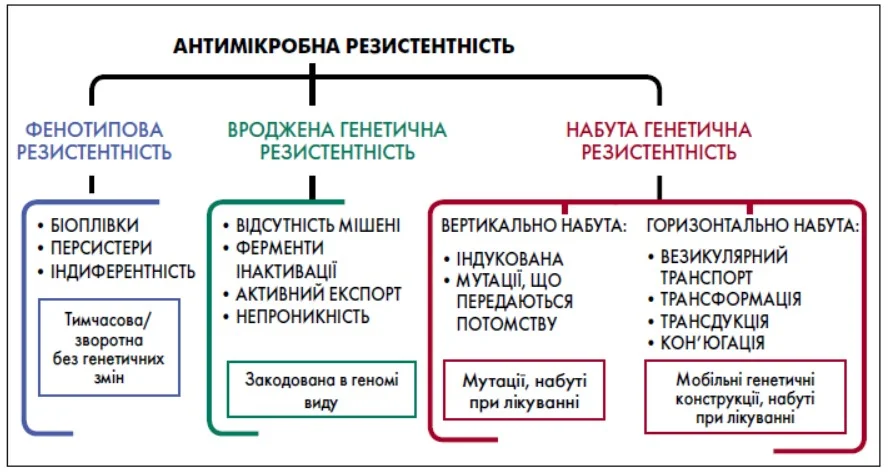
Рис.1. Антимікробна резистентність
Є комплексним біологічним явищем, що включає в себе як механізми, не закодовані в геномі безпосередньо, але які реалізуються через поведінку та життєвий цикл бактерії (фенотипова резистеність), так і генетично-опосередковані механізми, які або є частиною сталої генетичної програми виду (вроджена генетична резистентність), або набуваються протягом лікування пацієнта через як еволюційну адаптацію інфекційного штаму до антибіотика (вертикальна, індукована лікуванням генетична резистентність), так і через раптове запозичення генів резистентності у вигляді мобільної генетичної конструкції з госпітального середовища (горизонтально набута генетична резистентність) – так званого госпітального резистому, що підтримується в лікарні внаслідок недостатності заходів інфекційного контролю, з одного боку, та постійну присутність селективних умов, а саме високу концентрацію антибіотиків у лікарняному просторі – з іншого боку.
Фенотипова антибіотикорезистентність
Роль біоплівок
Ключовим компонентом АМР є фенотипова резистентність, основу якої становить утворення біоплівок. У реальних умовах бактерії переважно існують у вигляді структурованих мікробних спільнот, вбудованих у позаклітинний матрикс, що формуються на межі середовищ. Ці структуровані спільноти, або біоплівки, є базовою формою існування бактерій. Така організація обмежує дію антибіотиків та імунних механізмів і сприяє швидкому формуванню толерантних фенотипів, що має ключове значення для патогенезу інфекцій.
Принципово важливо відрізняти природні біоплівки від агарових колоній, які є штучною агрегацією клітин і не відтворюють структурної та функціональної організації бактеріальних спільнот in vivo; тому результати агаризованих моделей мають обмежену клінічну релевантність [2].
Додатковим ускладнюючим чинником є участь імунної системи макроорганізму, умови взаємодії з якою не відтворюються в агаризованих моделях (рис. 2, 3). Водночас навіть без імунного впливу організовані бактеріальні спільноти самі по собі суттєво обмежують ефективність антибіотикотерапії [3]. Зокрема, колістин і азтреонам на рівні вище мінімальної інгібуючої концентрації (МІК) не забезпечували повної елімінації бактерій: біоплівки зберігали метаболічну активність або зазнавали лише часткових змін.
 Рис. 2. Цитологічні картини гострого інфекційного полімікробного мікроландшафту на прикладі гострої стадії пародонтиту
Рис. 2. Цитологічні картини гострого інфекційного полімікробного мікроландшафту на прикладі гострої стадії пародонтиту
Інфекційний процес завжди тісно асоційований з імунною відповіддю пацієнта. При гострому інфекційному процесі запалення максимально інтенсивне, з високими рівнями прозапальних цитокінів (інтерлейкіни 6, 12, фактор некрозу пухлини та ін.), активним НЕТозом та М1-поляризацією макрофагів. На зображенні – під’ясенні мікроландшафти при пародонтиті, конфокальна лазерна скануюча мікроскопія (КЛСМ), забарвлення загальної ДНК бромідом етидію та йодидом пропідію (червоний сигнал) і клітини Candida spp., забарвлені калькофлуором (синій канал). Мікроландшафт демонструє велику кількість екзоклітинної ДНК (червоний колір), вірогідно, НЕТозного походження з невеликою кількістю адгезованих до неї клітин Candida (синій колір). Осі показують 90 мкм (адаптовано з Slobodianyk-Kolomoiets M., Khlebas S., Mazur I., Rudnieva K., Potochilova V., Iungin O., Kamyshnyi O., Kamyshna I., Potters G., Spiers A.J., Moshynets O. Extracellular host DNA contributes to pathogenic biofilm formation during periodontitis. Front Cell Infect Microbiol. 2024 May 8;14:1374817. doi: 10.3389/fcimb.2024.1374817).
 Рис. 3. Мікроландшафт при хронічному запаленні, під час якого імунна система намагається максимально обмежити вогнище запалення, на прикладі інфекційного ендокардиту
Рис. 3. Мікроландшафт при хронічному запаленні, під час якого імунна система намагається максимально обмежити вогнище запалення, на прикладі інфекційного ендокардиту
А. Гістологія гранульоми в реконструйованому з перикарду клапані аорти, КЛСМ, забарвлення ДНК бромідом етидію (червоний сигнал) і полісахаридів калькофлуором (синій канал). Гранульома (червоний сигнал) сформувалася навколо інфекційного вогнища (біоплівка, вкрита екзополісахаридами, які захищають її від фагоцитозу – синій колір).
Б. Сагітальний оптичний зріз із 3D КЛСМ-зображення, реконструйованого з А. Помітні окремі нитки червоної екзоклітинної ДНК, або еДНК (жовті стрілки) – НЕТозні екзоклітинні пастки, які обмежують розповсюдження інфекції на інші ділянки епітелію, зокрема зв’язуючи планктонні форми бактерій, що відшаровуються від біоплівки (отримано у співпраці з Колтуновою Г.Б. і Крикуновим О.А., відділення хірургічного лікування інфекційного ендокардиту Національного інституту серцево-судинної хірургії імeні М.М. Амосова).
Ще одним чинником, який знижує ефективність антибіотикотерапії, є здатність компонентів біоплівкового матриксу, зокрема позаклітинної ДНК, зв’язувати антибіотики. Відомо, що деякі групи препаратів, зокрема аміноглікозиди, можуть сорбуватися на ДНК внаслідок електростатичних взаємодій, що обмежує їх проникнення вглиб біоплівки та зменшує доступність для бактеріальних клітин.
Оскільки біоплівка є базовою формою існування бактерій, лікування інфекцій у тканинах потребує передусім фізичного усунення цього субстрату. Без санації вогнища антибіотикотерапія є неефективною. Клінічна тактика має включати дренування абсцесів, видалення інфікованих або некротизованих тканин і зменшення в’язкості секретів; за глибоких інфекцій (панкреатит, ендокардит) хірургічне втручання часто є критично необхідним.
Особливо складними є біоплівко-асоційовані інфекції на інертних поверхнях (катетери, імплантати), де антибіотикотерапія без видалення стороннього тіла не забезпечує ефективного лікування і може розглядатися лише як допоміжний метод.
Окремої уваги потребує питання оцінки синергічної дії антибіотиків. Використання агар-дифузійних методів для цієї мети не відображає фізіологічну реальність інфекційного процесу, асоційованого з біоплівковим ростом. Відмінності у фізіологічному стані бактерій, особливостях фармакокінетики та фармакодинаміки препаратів, а також відсутність валідованих стандартів інтерпретації призводять до того, що результати таких тестів не корелюють із реальною ефективністю комбінованої антибіотикотерапії в клінічних умовах [4]. Більш детально ми розглядали питання доказовості синергідних тестів раніше [5].
Значення персисторних клітин
Важливим компонентом фенотипової АМР є персисторні клітини – невелика за чисельністю (≈1%) субпопуляція бактерій, що формується до початку антибіотикотерапії та не пов’язана з генетичними мутаціями. Вони характеризуються глибоко зниженою метаболічною активністю, що зумовлює високу толерантність до антибіотиків, ефективних переважно щодо активно проліферуючих клітин. Персистори можуть існувати як у планктонній формі, так і в біоплівках, однак саме останнє середовище є основною нішею їх формування та збереження.
Формування персисторних субпопуляцій – регульована відповідь на стресові чинники, зокрема субінгібуючі концентрації антибіотиків та дефіцит поживних ресурсів, і супроводжується переходом частини клітин у стан глибокого метаболічного спокою. У клінічних умовах це призводить до елімінації основної маси бактерій при збереженні персисторів, які підтримують хронізацію інфекції та виступають ключовим джерелом гетерорезистентності й проміжною ланкою між фенотиповою толерантністю та формуванням стабільної генетичної АМР.
З клінічної точки зору терапевтично досяжні концентрації антибіотиків мають бути активними не лише щодо проліферуючих клітин, а й щодо персисторних субпопуляцій, які є найменш доступними для стандартної монотерапії та зумовлюють потребу в призначенні комбінованих схем.
Прикладом клінічно значущої персистенції є рецидивуючі інфекції сечових шляхів, асоційовані з Klebsiella pneumoniae. У таких випадках персистування збудника спричинене не лише біоплівковими механізмами, а й альтернативною адаптивною стратегією, пов’язаною зі зменшенням або втратою капсульного шару та проникненням бактерій у клітини уротелію. У внутрішньоклітинних вакуолях бактерії переходять у персистуючий стан, що робить їх практично недосяжними для антибіотиків навіть у високих дозах.
Саме цей механізм пояснює високу частоту клінічних рецидивів і вказує на обмеженість сучасних терапевтичних підходів [6]. Допоміжні стратегії, зокрема застосування D-маннози для інгібування адгезії ентеробактерій до уротелію, мають певне клінічне підґрунтя, однак наразі не інтегровані у стандартизовані рекомендації.
Індиферентність до антибіотиків
Ще одним важливим, але часто недооціненим проявом фенотипової АМР є індиферентність – тимчасова функціональна нечутливість бактеріальної популяції до антибіотиків, не пов’язана з генетичними механізмами резистентності. Вона зумовлена фізіологічним станом бактерій і може спостерігатися навіть за формально збереженої генетичної чутливості. Показовим прикладом є колістин щодо K. pneumoniae: за умов чутливості in vitro препарат може частково впливати на біоплівкові структури, тоді як планктонна популяція залишається практично нечутливою до його дії. Цей феномен частково пояснює низьку клінічну ефективність колістину при гострих системних інфекціях, зокрема сепсисі, попри підтверджену чутливість збудника.
Вроджена генетична резистентність
На відміну від фенотипових форм, вроджена генетична резистентність є найменш проблемною з клінічної точки зору, оскільки вона стабільна, добре вивчена та зумовлена видовими особливостями бактерій. Її механізми включають відсутність мішеней для дії антибіотиків, низьку проникність клітинної стінки, активний ефлюкс і наявність вроджених ферментів інактивації.
Класичним прикладом цього є природна резистентність Pseudomonas aeruginosa до цефтріаксону, пов’язана з активним ефлюксом і продукцією хромосомної β-лактамази AmpC; у таких випадках відсутність клінічного ефекту є очікуваною і не має адаптивної природи.
Дані про вроджену резистентність систематизовані в рекомендаціях Європейського комітету з тестування антимікробної чутливості (EUCAST), які слугують валідованою основою для інтерпретації антибіотикограм в Україні та дозволяють заздалегідь виключати неефективні терапевтичні опції [7].
Набута генетична резистентність
Набута генетична АМР є центральною проблемою сучасної антимікробної терапії та основою глобальної кризи резистентності. На відміну від фенотипової АМР, яка часто має зворотний характер і пов’язана з фізіологічним станом популяції, генетично набута стійкість призводить до формування стабільних, клінічно значущих резистентних клонів. Принципово важливо розрізняти вертикально та горизонтально набуту генетичну резистентність.
Вертикальна резистентність формується безпосередньо під час лікування пацієнта внаслідок мутацій у межах однієї бактеріальної популяції під селективним тиском антибіотиків. Класичним прикладом є розвиток резистентності Staphylococcus aureus до лінезоліду за умови некоректної терапії, що призводить до появи резистентних субпопуляцій у джерелі інфекції. Така резистентність часто має високу біологічну «ціну», тому після припинення антибіотикотерапії резистентні клони зазвичай витісняються чутливими, що робить цей тип АМР відносно передбачуваним і потенційно контрольованим.
Натомість горизонтально набута генетична резистентність є найбільш загрозливою, особливо серед грамнегативних бактерій. Вона формується внаслідок циркуляції генів резистентності в госпітальному середовищі та лежить в основі появи карбапенем-резистентних грамнегативних патогенів – однієї з найсерйозніших терапевтичних проблем сучасної медицини.
Карбапенеми належать до найефективніших антибіотиків, поєднуючи широкий спектр дії, виражену бактерицидність, високу біодоступність і відносно низьку токсичність. Втрата їх клінічної ефективності має критичні наслідки для лікування тяжких інфекцій.
Резистентність до карбапенемів у грамнегативних бактерій формується внаслідок поєднання кількох механізмів, зокрема [8]:
- зниження проникності зовнішньої мембрани;
- активації ефлюкс-систем;
- продукції β-лактамаз, включно з карбапенемазами;
- супутніх механізмів резистентності до інших класів антибіотиків.
Саме ця багаторівнева комбінація робить карбапенем-резистентні штами практично нечутливими до стандартних схем лікування та змушує клініцистів вдаватися до емпіричних або експериментальних підходів.
Еволюція антибіотикорезистентності під час війни в Україні
В Україні АМР сформувалася задовго до 2022 року, однак повномасштабна війна різко прискорила її поширення. Перевантаження стаціонарів, застосування емпіричної антибіотикотерапії, порушення інфекційного контролю та обмежена діагностика створили умови для швидкої експансії резистентних грамнегативних патогенів. Динаміка змін рівнів резистентності основних груп збудників до ключових класів антибіотиків наведена на рис. 4.
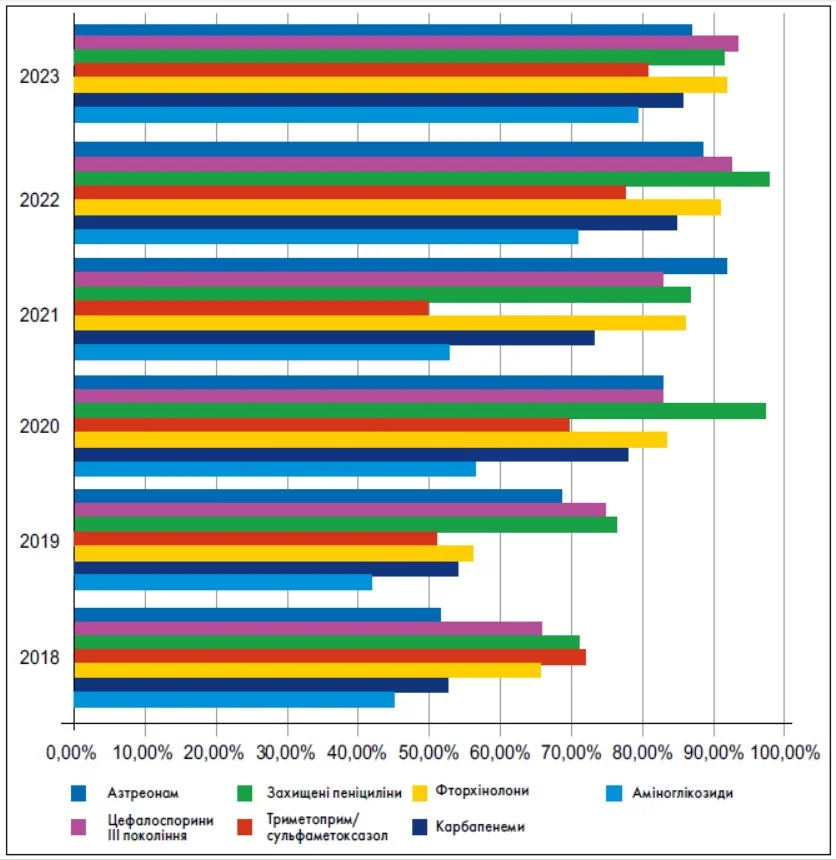
Рис. 4. Тенденція зростання резистентності до найпоширеніших антибактеріальних препаратів протягом 2018-2023 рр. на прикладі Київської обласної клінічної лікарні
(за даними Руднєвої К.Л., керівника відділу інфекційного контролю, лікаря-епідеміолога, лікаря-бактеріолога бактеріологічного відділу Центру лабораторних досліджень КЛ «Феофанія» ДУС).
Таким чином, починаючи з 2018 року рівні АМР до цефалоспоринів III покоління, фторхінолонів, захищених пеніцилінів та аміноглікозидів щороку зростали, а до 2023 року в стаціонарних умовах для більшості цих класів наблизилися або перевищили 90%. Карбапенеми, які традиційно розглядалися як антибіотики резерву, також поступово втрачали ефективність, що відображає накопичення мультимеханізмової резистентності серед грамнегативних бактерій.
Ці локальні спостереження узгоджуються з результатами багатоцентрового дослідження, яке охопило понад 6 800 клінічних ізолятів із великих українських стаціонарів [9]. Автори продемонстрували приблизно рівне співвідношення грампозитивної та грамнегативної флори, однак із принципово різною чутливістю до антибіотикотерапії. Грампозитивні збудники зберігали високу чутливість до ключових препаратів, зокрема майже 100% – до ванкоміцину та лінезоліду. Натомість грамнегативні бактерії характеризувалися критично низькою чутливістю до основних терапевтичних опцій.
Чутливість грамнегативних ізолятів до меропенему в окремих центрах становила лише 15-30%, до іміпенему – подекуди була близькою до нуля. Навіть сучасні комбінації, зокрема цефтазидим/авібактам, демонстрували обмежену ефективність, тоді як чутливість до азтреонаму коливалася в межах 20-30%.
Ключова причина такої різниці між грампозитивними та грамнегативними бактеріями полягає в особливостях клітинної оболонки останніх: вони поєднують периплазматичну локалізацію β-лактамаз і карбапенемаз, активні ефлюкс-системи та регульовану проникність поринів. Резистентність до карбапенемів формується внаслідок комбінації цих механізмів.
Важливо, що українські карбапенем-резистентні грамнегативні штами суттєво відрізняються від ізолятів Західної Європи та США за механізмами резистентності та роллю ефлюкс-систем. У зв’язку з цим терапевтичні схеми, ефективні в західних країнах (зокрема, цефтазидим/авібактам з азтреонамом), в українських клінічних умовах часто не дають очікуваного ефекту. За відсутності національного молекулярного моніторингу АМР ці відмінності залишаються недостатньо вивченими, особливо в умовах війни.
Одним із небагатьох системних джерел даних про молекулярні механізми карбапенем-резистентності українських штамів у перші роки повномасштабного вторгнення є дослідження польських колег [10], виконане на базі Національного референс-центру Польщі, ключовою цінністю якого є аналіз генетичної структури АМР.
У дослідженні проаналізовано 65 XDR/PDR ізолятів K. pneumoniae, отриманих від українських пацієнтів у 2022-2023 роках, які були евакуйовані або переведені на лікування до Польщі (реальна клінічна популяція госпітальних інфекцій).
Було показано домінування ферментативних механізмів карбапенем-резистентності з переважанням метало-β-лактамаз. Найпоширенішою карбапенемазою була NDM (New Delhi металo-β-лактамази), яку виявлено у 44,6% ізолятів. Друге місце посідали KPC (карбапенемази K. pneumoniae, 24,6%), далі – OXA‑48-like (12,3%) та VIM (1,5%). Особливо тривожним є те, що 17% ізолятів одночасно несли два типи карбапенемаз, найчастіше – комбінацію NDM + OXA‑48-like, що практично виключає ефективність монотерапії.
Домінування NDM формує так званий азійський тип резистентності, відмінний від західноєвропейського та північноамериканського профілю з переважанням KPC. Для України, розташованої на транзитному шляху між Азією та Європою, така структура є очікуваною, але клінічно несприятливою.
З метою оцінки подальшої еволюції механізмів резистентності у 2024 році було проаналізовано близько 100 карбапенем-резистентних ізолятів із Києва, Харкова та Кривого Рогу, що належали до трьох основних груп грамнегативних бактерій: Enterobacterales, Acinetobacter baumannii та Pseudomonas aeruginosa [11].
Принципово важливим спостереженням стало те, що наявність карбапенемази не була універсальною ознакою резистентності: у P. aeruginosa близько третини ізолятів не продукували карбапенемази, що вказує на провідну роль неферментативних механізмів – зниження проникності та активації ефлюкс-систем. Для Enterobacterales та A. baumannii ферментативні механізми залишалися більш типовими, однак відзначалися значною гетерогенністю.
Порівняльний аналіз фенотипової чутливості карбапенем-резистентних Enterobacterales у 2020, 2023 і 2024 роках виявив стійку негативну динаміку для більшості терапевтичних опцій, зокрема подальше зниження чутливості до амікацину та колістину у 2024 році, що свідчить про втрату одних із небагатьох залишкових альтернатив.
Особливе занепокоєння викликає зниження чутливості до цефтазидиму/авібактаму: у 2024 році чутливими залишалися лише близько третини ізолятів. Ще більш показовою є динаміка цефідероколу – падіння чутливості зі 100% у 2020 році до близько 50% у 2024 році, що за умов обмеженої доступності препарату в Україні вказує на формування перехресної резистентності.
Виявлення карбапенем-резистентних Enterobacterales без карбапенемаз свідчить про зміщення домінуючих механізмів резистентності від ферментативних до неферментативних – зниження проникності та посилення ефлюксу. Загалом, це відображає подальшу еволюцію карбапенем-резистентності в Україні та формування клінічних сценаріїв без ефективних і доступних терапевтичних опцій для тяжких грамнегативних інфекцій.
Нормативні рекомендації та їх обмеження в українських реаліях
Чинні нормативні рекомендації, зокрема наказ МОЗ України № 1513 від 23.08.2023 р., попри декларовану відповідність принципам доказової медицини, недостатньо враховують клінічні реалії України. Запропоновані схеми переважно базуються на західних протоколах без адаптації до локальної епідеміології, структури резистентності та доступності препаратів. Частина антибіотиків (зокрема, цефтолозан/тазобактам) відсутні або лімітовано доступні, а інші комбінації є економічно недосяжними й не покриваються Національною службою здоров’я України, що обмежує їх практичне застосування при карбапенем-резистентних грамнегативних інфекціях.
Водночас в Україні відсутня власна доказова база ефективності рекомендованих схем через обмежений доступ до препаратів і відсутність системного збору клінічних даних, унаслідок чого наявні спостереження залишаються фрагментарними.
Проблема лікування карбапенем-стійких грамнегативних інфекцій: світовий досвід і власні дослідження
За відсутності власних даних доцільно звертатися до міжнародного досвіду з регіонів із подібною структурою механізмів резистентності. На Європейському конгресі клінічної мікробіології та інфекційних захворювань (ECCMID) у 2024 році було представлено результати досліджень із Сінгапуру, де бактерицидна активність комбінації цефтазидиму/авібактаму в поєднанні з азтреонамом становила близько 46%, що свідчить про обмежену ефективність цієї схеми навіть у добре контрольованих умовах (Jocelyn T., National University Health System, Singapore).
Подібні результати отримано й в Індії – регіоні з високою поширеністю метало-β-лактамаз [12]. У ретроспективному когортному дослідженні CAPRІ (106 пацієнтів із карбапенем-резистентними інфекціями, спричиненими K. pneumoniae) летальність у групі колістину становила 57%, у групі цефтазидиму/авібактаму – 29%, а при застосуванні комбінації цефтазидим/авібактам + азтреонам – 36%, без статистично значущої різниці між групами. Отже, додавання азтреонаму не забезпечувало клінічно значущого покращення результатів.
У власних дослідженнях ми розглянули карбапенем-резистентність з альтернативної позиції, зосередившися на ролі біоплівок у персистенції інфекції. У значної частки клінічних ізолятів зберігалася чутливість лише до колістину, однак його клінічна ефективність є обмеженою, зокрема через погану біодоступність у біоплівкові структури. Це дозволило висунути гіпотезу, що підвищення ефективності терапії можливе шляхом одночасного порушення біоплівки.
Враховуючи відомий антибіоплівковий ефект азитроміцину при інфекціях, спричинених P. aeruginosa, і спільність регуляторних механізмів біоплівкоутворення у P. aeruginosa та K. pneumoniae, було оцінено антибіоплівкову активність макролідів щодо останнього патогена. Із протестованих препаратів лише азитроміцин і джозаміцин продемонстрували виражене пригнічення біоплівки; з огляду на наявність парентеральної форми подальші дослідження зосередилися на азитроміцині [13]. Він виявився ефективнішим за колістин як у планктонних, так і в біоплівкових формах, тоді як колістин у монорежимі мав мінімальний або негативний ефект. Комбінація азитроміцину з колістином забезпечувала виражену синергію та повне пригнічення інфекційного процесу в розробленій in vitro моделі, тоді як монотерапія колістином була неефективною [14].
Отримані результати підтверджують доцільність таргетування біоплівки як окремої фармакологічної мішені та демонструють потенціал макролідів як засобів підвищення ефективності терапії карбапенем-резистентних інфекцій.
Азитроміцин-індукована колатеральна чутливість до деяких b-лактамів є новим біологічним феноменом, який було описано при спостереженні за терапією карбапенем-резистентних грамнегативних інфекцій комбінаціями антибіотиків з азитроміцином [15]. З’ясувалося, що за умов високодозової терапії азитроміцином, а саме 1500 мг на добу, що розділені на три парентеральні введення, спостерігалася дерепресія імунної відповіді, що суттєво покращувало стан пацієнта, підсилювало його опір інфекції, знижувало запальне навантаження в умовах екзотоксин-індукованого цитокінового шторму та відбувалася зміна чутливості збудника до деяких β-лактамних антибіотиків, зокрема до меропенему в меропенем-резистентних ентеробактерій. Вірогідно, це пов’язано з еволюційною адаптацією грамнегативного штаму до азитроміцину за фізіологічних умов, оскільки показано, що за таких умов він проявляє набагато більшу бактерицидність, ніж у традиційних мікробіологічних тестах [16]. Така висока бактерицидність азитроміцину призводить до селекції мутацій у популяції збудника, що підвищують толерантність до азитроміцину, але в той же час корелюють із появою чутливості до меропенему. Швидше за все, це відбувається через перебудову клітинної стінки, яка втрачає механізми резистентності до меропенему попри генетичну резистентність до нього.
Описана колатеральна чутливість до меропенему відкриває вікно терапевтичних можливостей для лікування таких меропенем-резистентних інфекцій, що було використано в успішному лікуванні різноманітних клінічніх випадків, зорема сечових інфекцій, сепсису, ранових інфекцій, остеомієліту та ін. Для цього терапія азитроміцином у дозуванні 1500 мг на добу комбінувалася з меропенемом у добовій дозі 6000 мг. Важливо наголосити, що така колатеральна чутливість не може бути виявлена у класичних диск-дифузійних тестах через невідповідність форми існування інфекційного ізолята (колонія на агарі фізіологічно не відповідає біоплівці цього штаму за патофізіологічних умов) і мікрооточення – відсутність компонентів імунної системи пацієнта, білкового середовища тощо.
Висновки
АМР грамнегативних бактерій в Україні є складною багатофакторною проблемою, що формується внаслідок поєднання генетичних і фенотипових механізмів, зокрема утворення біоплівок, появи персисторних субпопуляцій та активації альтернативних шляхів захисту від дії антимікробних препаратів. Отримані дані свідчать, що ефективність антибіотикотерапії не може бути адекватно оцінена виключно на підставі стандартної антибіотикограми, оскільки фенотипові форми стійкості суттєво впливають на клінічний перебіг інфекційного процесу.
Аналіз вітчизняних і міжнародних досліджень демонструє, що структура механізмів карбапенем-резистентності в Україні має виражені регіональні особливості й за своїми характеристиками наближається до азійського типу. Це суттєво обмежує можливість прямої екстраполяції західних терапевтичних протоколів без попередньої локальної валідації. Формально затверджені національні рекомендації не завжди враховують реальну епідеміологічну ситуацію, доступність антибактеріальних препаратів та економічні обмеження, що знижує їхню практичну цінність у лікуванні пацієнтів із карбапенем-резистентними інфекціями.
Світовий досвід застосування комбінованих схем терапії, зокрема комбінації цефтазидиму/авібактаму з азтреонамом, свідчить про обмежену й неоднозначну клінічну ефективність, особливо в регіонах із високою поширеністю метало-β-лактамаз. Це підкреслює необхідність обережного використання таких підходів за відсутності власної доказової бази. У цьому контексті перспективними видаються альтернативні стратегії, спрямовані на порушення структури біоплівки та підвищення доступності антибіотика у вогнищі інфекції, а також нещодавно описане явище азитроміцин-індукованої колатеральної чутливості, коли високодозова терапія азитроміцином індукує колатеральну чутливість карбапенем-резистентних грамнегативних бактерій до окремих β-лактамів, зокрема меропенему. Ймовірним механізмом є адаптивна перебудова клітинної стінки під впливом підвищеної бактерицидності азитроміцину за фізіологічних умов. Комбінація азитроміцину з меропенемом відкриває нові терапевтичні можливості, які не виявляються стандартними лабораторними тестами.
Таким чином, ефективне подолання проблеми антимікробної резистентності в Україні потребує розвитку національної системи молекулярного та фенотипового моніторингу, проведення власних клінічних і молекулярно-генетичних досліджень, а також перегляду терапевтичних підходів з урахуванням локальних епідеміологічних і біологічних особливостей збудників.
Література
- Olivares J., Bernardini A., Garcia-Leon G., Corona F., B Sanchez M., Martinez J.L. The intrinsic resistome of bacterial pathogens. Front Microbiol. 2013 Apr 30;4:103. doi: 10.3389/fmicb.2013.00103.
- Cornforth D.M., Dees J..L, Ibberson C.B., Huse H.K., Mathiesen I.H., Kirketerp-Mшller K., Wolcott R.D., Rumbaugh K.P., Bjarnsholt T., Whiteley M. Pseudomonas aeruginosa transcriptome during human infection. Proc Natl Acad Sci U S A. 2018 May 29;115(22):E5125-E5134. doi: 10.1073/pnas.1717525115.
- Mьsken M, Klimmek K, Sauer-Heilborn A, Donnert M, Sedlacek L, Suerbaum S, Hдussler S. Towards individualized diagnostics of biofilm-associated infections: a case study. NPJ Biofilms Microbiomes. 2017 Sep 28;3:22. doi: 10.1038/s41522-017-0030-5.
- Moshynets O.V., Baranovskyi T.P., Cameron S., Iungin O.S., Pokholenko I., Jerdan R., Kamyshnyi A., Krikunov A..A., Potochilova VV., Rudnieva K.L., Spiers A.J. Azithromycin possesses biofilm-inhibitory activity and potentiates non-bactericidal colistin methanesulfonate (CMS) and polymyxin B against Klebsiella pneumonia. PLoS One. 2022 Jul 1;17(7):e0270983. doi: 10.1371/journal.pone.0270983.
- https://health-ua.com/anesteziologiya/mizdisciplinarni-problemi/80052-iak-ukrayinska-medicna-sistema-vidpovidaje-na-viklik-antibiotikorezistentnosti-ci-stanut-sinergidni-testi-risenniam-problemi.
- Ernst C.M., Braxton J.R., Rodriguez-Osorio C.A., Zagieboylo A.P., Li L., Pironti A., Manson A.L., Nair A.V., Benson M., Cummins K., Clatworthy A.E., Earl A.M., Cosimi L.A., Hung D.T. Adaptive evolution of virulence and persistence in carbapenem-resistant Klebsiella pneumoniae. Nat Med. 2020 May;26(5):705-711. doi: 10.1038/s41591-020-0825-4. Epub 2020 Apr 13. Erratum in: Nat Med. 2020 Aug;26(8):1307. doi: 10.1038/s41591-020-0974-5.
- https://www.eucast.org/expert_rules_and_expected_phenotypes.
- https://www.intechopen.com/chapters/70581.
- Uren H., Aliieva N., Matolinets N., Hemsley C., Holcomb J.B., Samchuk O., Herych H. Cumulative antibiogram results of over 6800 diagnostic isolates from the First Lviv Territorial Medical Union, Ukraine. Lancet Infect Dis. 2025 Apr;25(4):e200-e201. doi: 10.1016/S1473-3099(25)00116-1.
- Biedrzycka M., Izdebski R., Hryniewicz W., Gniadkowski M., Żabicka D. Carbapenemase-Producing Enterobacterales from Patients Arriving from Ukraine in Poland, March 2022-February 2023. Infect Dis Ther. 2025 Feb;14(2):401-419. doi: 10.1007/s40121-024-01097-9.
- Pallett S.J.C., Morkowska A., Woolley S.D., Potochilova V.V., Rudnieva K.L., Iungin O.S., Sgro V., Boyd S.E., Reece N., Lambert Z.L., Tan N.K., Mughal N., Moshynets O.V., Moore L.S.P., O’Shea M.K. Evolving antimicrobial resistance of extensively drug-resistant Gram-negative severe infections associated with conflict wounds in Ukraine: an observational study. Lancet Reg Health Eur. 2025 Mar 20;52:101274. doi: 10.1016/j.lanepe.2025.101274.
- Sree R.A., Gupta A., Gupta N., Veturi S., Reddy L.S.K., Begum M., Shravani E., Challa H.R., Reddy S.S., Singamsetty A., Arumilli M., Reddy P.N., Tirlangi P.K. Ceftazidime-avibactam alone or in combination with Aztreonam versus Polymyxins in the management of carbapenem-Resistant Klebsiella pneumoniae nosocomial Infections (CAPRI study): a retrospective cohort study from South India. Infection. 2024 Apr;52(2):429-437. doi: 10.1007/s15010-023-02094-9.
- Moshynets O.V., Baranovskyi T.P., Cameron S., Iungin O.S., Pokholenko I., Jerdan R., Kamyshnyi A.., Krikunov AA., Potochilova V.V., Rudnieva K.L., Spiers A.J. Azithromycin possesses biofilm-inhibitory activity and potentiates non-bactericidal colistin methanesulfonate (CMS) and polymyxin B against Klebsiella pneumonia. PLoS One. 2022 Jul 1;17(7):e0270983. doi: 10.1371/journal.pone.0270983.
- Moshynets O.V., Baranovskyi T.P., Iungin O.S., Krikunov A.A., Potochilova V.V., Rudnieva K.L., Potters G., Pokholenko I. Therapeutic Potential of an Azithromycin-Colistin Combination against XDR K. pneumoniae in a 3D Collagen-Based In Vitro Wound Model of a Biofilm Infection. Antibiotics (Basel). 2023 Feb 1;12(2):293. doi: 10.3390/antibiotics12020293.
- Kryzhevskyi V., Strokous V., Lifshyts Y., Rybianets Y., Oberniak A., Krikunov A., Iungin O., Potochilova V., Rudnieva K., Petakh P., Kamyshnyi A., Moshynets O. Case report: Azithromycin-meropenem combination therapy as a low-cost approach to combat PDR gram-negative infections of war wounds in Ukraine. Front Med (Lausanne). 2023 Sep 22;10:1264492. doi: 10.3389/fmed.2023.1264492.
- Buyck J.M., Plйsiat P., Traore H., Vanderbist F., Tulkens P.M., van Bambeke F. Increased susceptibility of Pseudomonas aeruginosa to macrolides and ketolides in eukaryotic cell culture media and biological fluids due to decreased expression of oprM and increased outer-membrane permeability. Clin Infect Dis. 2012 Aug;55(4):534-42. doi: 10.1093/cid/cis473.
Підготувала Ганна Кирпач
Тематичний номер «Хірургія. Ортопедія. Травматологія. Інтенсивна терапія» № 5 (67), 2025 р.