


27 грудня, 2018
Танці очей
За матеріалами X Міжнародної конференції для неврологів, психіатрів, психотерапевтів, лікарів суміжних спеціальностей «Нейросимпозіум» (11-13 вересня 2018 року, м. Одеса)
 На конференції першу частину циклу лекцій про сакадні системи загалом, порушення сакад та особливості клінічної картини, фармакотерапію осцилопсії тощо, а саме власне бачення цих процесів представив завідувач кафедри неврології, нейрохірургії та психології Ужгородського національного університету, доктор медичних наук, професор Михайло Михайлович Орос.
На конференції першу частину циклу лекцій про сакадні системи загалом, порушення сакад та особливості клінічної картини, фармакотерапію осцилопсії тощо, а саме власне бачення цих процесів представив завідувач кафедри неврології, нейрохірургії та психології Ужгородського національного університету, доктор медичних наук, професор Михайло Михайлович Орос.
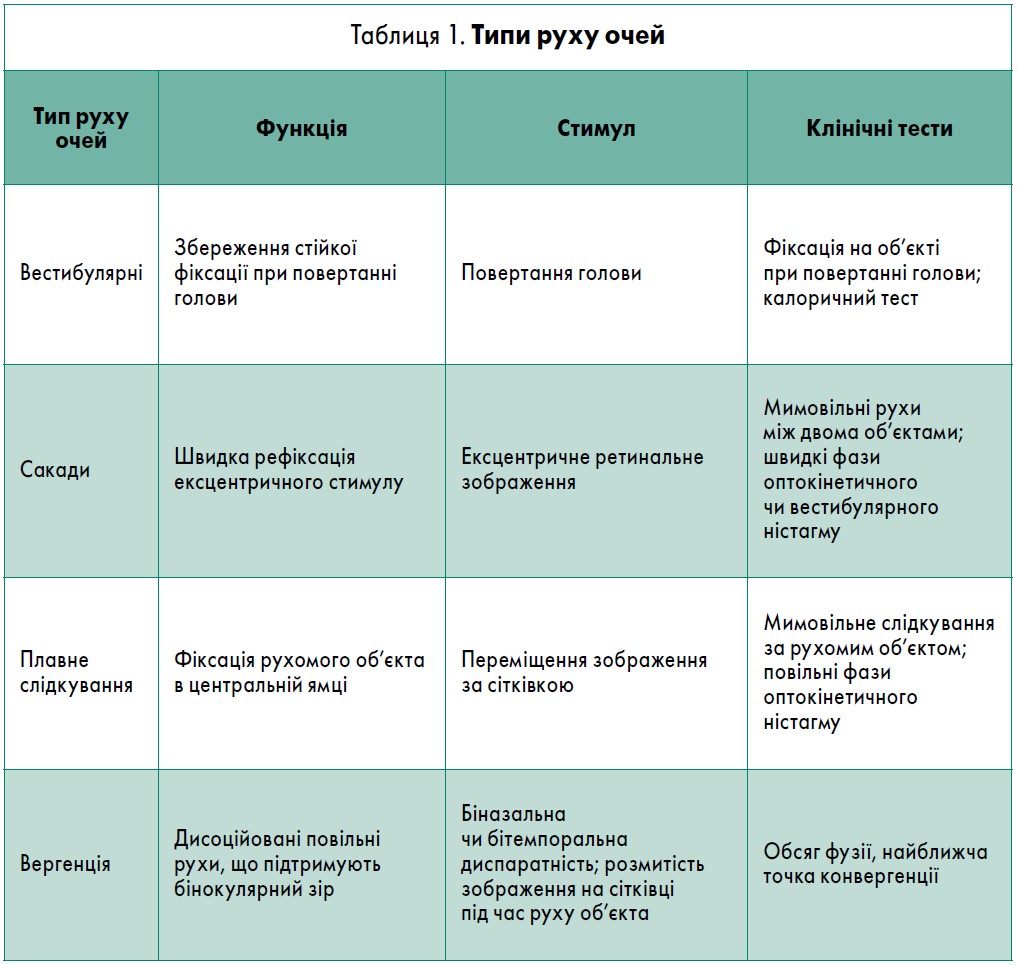
У своїй доповіді лектор доречно згадав слова засновника американської нейроофтальмології, доктора Девіда Зі (David Zee) про те, що ми всі живемо на краю стабільності з нашими сакадними системами. Сакадні рухи – це системи, що дають можливість бачити світ стабільно. Яку ж саме функцію виконують окорухові м’язи наших очей узагалі? Вони слугують для того, щоб зображення потрапило в центр найкращого зору. Завдяки окоруховим м’язам ми рухаємо очима, бачимо в різні боки. Вони пов’язані з різними положеннями нашого тіла, голови й тим, що відбувається навколо нас. Типи руху очей, їхні функції та стимули наведені в таблиці 1.
Окулоцефалічний рефлекс – це рухомі реакції очей на подразнення вестибулярного апарату, опосередковані шляхами, що проходять крізь стовбур мозку від вестибулярних ядер довгастого мозку до ядер відвідного й окорухового нервів. Окорухову функцію, яку виконують групи клітин мозочка, ядра під’язикового нерва та медіального вестибулярного ядра, називають нейронним інтегратором. Щоб рухати очима з постійною швидкістю й утримувати в ексцентричній позиції погляду, два нервові сигнали мають протидіяти еластичній тязі, що прагне повернути очі в положення «спокою». Такі сигнали називаються тонічними та фазовими компонентами. Механізм найменших змін тонічного компонента, що регулюються премоторним нервовим сигналом, і є нейроінтегратором. Нейрофізіологічні дані показують, що команда позиції (наприклад, для сакад, інтервалу) генерується з команди швидкості (приміром, для сакад, імпульсу) математичним процесом інтеграції стосовно часу. Нейронна мережа інтегрує в цьому математичному сенсі сигнали, що кодують швидкість, у сигнали, що кодують положення.
Система сакад та її функціонування
Сакади (в перекладі з давньофранцузької – «хлопок вітрила») – швидкі, чітко узгоджені рухи очей, що відбуваються одночасно та в одному напрямку. Вони можуть виникати мимовільно, рефлекторно чи бути частиною швидкофазових рухів під час ністагму. Сакади генерує стовбур мозку. Черв’як мозочка, ядро намету мозочка та флокулус беруть участь у калібруванні та модуляції сакадних реакцій. Нервові вибухові нейрони (в межах мосту й середнього мозку) виділяють від 10 до 12 мс в очікуванні сакади та випускають до мотонейронів іпсилатерального ядра відповідного нерва. Інгібіторні нейрони випускають до шостого ядра C/L.
Універсальні паузові нейрони розподіляються по всьому стовбуру головного мозку й чинять тонічний інгібувальний ефект. Їхня дія спрямована насамперед на запобігання небажаним і нав’язливим сакадам. Універсальні паузові нейрони закінчують активність приблизно за 15 мс до сакади, що дає змогу нервовим вибуховим нейронам активізуватися. Також інгібіторні вибухові нейрони пригнічують активацію антагоністичних екстраокулярних м’язів. Ядро відповідного нерва є горизонтальним центром погляду й отримує команди від усіх центрів, що забезпечують рухи очей.
Сакадний контроль
Кора головного мозку бере участь у контролі всіх класів руху очей. Узагалі рефлекторні збудливоспрямовані рухи очей походять із задньої частини мозку, тоді як добровільні виникають із фронтальних ділянок. Фронтальна кора містить декілька ділянок, відповідальних за ініціювання горизонтальних сакад. Вони включають: фронтальне окорухове поле (FEFs), додаткове окорухове поле (SEFs) та дорсолатеральну префронтальну кору (DLPC).
Нейрони FEFs виділяються для сакади довільної, сакади, що керується пам’яттю, та рухів вергенції. Нейрони SEFs беруть участь у вивчених моделях поведінки окулярних рухів. DLPC контролює заплановані сакади до згаданих цілей. Тім’яні окорухові поля (PEFs) спрямовуються до FEFs і залучені у вивчення візуальних сцен та ініціювання рефлексивних візуально керованих сакад. Що стосується FEFs та PEFs, то вони значною мірою взаємопов’язані між собою. Скронево-тім’яно-потиличне (TPO) з’єднання займається сприйняттям візуальних рухів та плавним переслідуванням. Воно також відіграє значну роль у візуальній фіксації та утриманні погляду.
Сакадний шлях
Контроль горизонтальних сакад у півкулі є контралатеральним (розташований на іншому боці). Сигнали, що виникають унаслідок FEF (переважно невізуально керовані сакади) та PEF (переважно візуально керовані сакади), спускаються до імпульсних клітин контралатеральної парамедіанної ретикулярної формації моста (PPRF). Один фронтальний шлях проектується безпосередньо на PPRF, а інший – через хвостату, чорну субстанцію та верхній пагорбок (SC), перш ніж потрапити до PPRF. Шляхи через базальні ганглії підтримують рівновагу між рефлексивними та цілеспрямованими добровільними сакадами й допомагають запобігти нав’язливим сакадам. PEF також реалізує проекти через SC до PPRF.
Основні напрямки руху та типи сакад
Фізіологічні сакади зникають при фіксації погляду, інтрузивні ж являють собою неадекватні сакадичні рухи, що не зникають при фіксації. Для виявлення останніх необхідний кількісний запис руху очей.
Інтрузивні сакади очей з інтерсакадним інтервалом (ІСІ) – пари маленьких горизонтальних сакад з амплітудою 0,5°, представлені квадратними хвилями-поштовхами, що проходять від та до серединної лінії. Повернення до точки фіксації відбувається при ІСІ 200 мс. У пацієнтів, частіше в старшому, а може бути в середньому віці, спостерігаються труднощі фіксації погляду (часто вона неможлива), на осцилопсію типово не скаржаться. Макросакади з ІСІ – інтрузивні сакади з осциляціями навколо серединної лінії, горизонтальні. Вони піднімаються вище точки фіксації, тобто спостерігається збільшення/зменшення амплітуди. Фіксація погляду неможлива (ІСІ – 200 мс). Такі сакади є симптоматичними та свідчать про осцилопсію. Тремтіння очей (флатер) без ІСІ – сакади пліч-о-пліч, із високою частотою та горизонтальними осциляціями, викликаються конвергенцією. Опсоклонус без ІСІ – інтрузивні сакади пліч-о-пліч у всіх напрямках, переривчасто-вибухові, дуже швидкі, з горизонтальним, вертикальним, ротаційним компонентами. Є типово симптоматичними та вказують на осцилопатію. Тремтіння очей та опсоклонус, можливо, виникають через нестабільність у біполярному нейроні пауз. Локалізуються в OPNs, ядрі намету й асоціюються з міоклонусом, атаксією, емоційною лабільністю.
Порушення сакад
Час ініціації сакади (латентність) перебуває під впливом предмету, уваги та рівня свідомості. Лобно-тім’яні ураження можуть спричинити набуття окулярної моторної апраксії з різко порушеною активацією вольових сакад, тоді виникають мимовільні сакади (наприклад, гемодинамічний інсульт). Парієтальна кора є посередником для сакад у напрямку нових візуальних цілей. Однобічні парієтальні ураження можуть спричинити гіпометричні контралатеральні сакади. Як правило, це помітніше в разі правого (недомінантного) ураження. Хвороба базальних ганглій може призвести до зростання затримок при утворенні сакад: більших – при хворобі Гентінгтона, менших – при хворобі Паркінсона. Розлади сакадної точності означають захворювання мозочка й зазвичай призводять до гіперметрії (перевищення сакад). Основні типи та причини порушень сакад наведені в таблиці 2.

Клінічна оцінка сакад
Щодо патології сакади можна поділити на: розлади ініціації (тривалість і латентність) та швидкості (повільні сакади), відсутні чи небажані сакади, порушення точності (гіпо-/гіперметричні сакади).
У клініці добровільні сакади оцінюються шляхом запиту пацієнта на рефіксацію між двома цілями (наприклад, пальцями екзаменатора), як правило, відокремлені на 30-40°. Звичайні рухи рефіксації мають виконуватися за допомогою однієї сакади або можуть понизити ціль і потребувати однієї чи двох розв’язувальних сакад для досягнення мети. Три чи більше рефіксаційні сакади вважаються гіпометричними та ненормальними, особливо якщо асиметричні. Гіперметричні сакади перевищують цілі та завжди аномальні, що часто вказує на дисфункцію мозочкової системи. Сакадна гіпометрія вказує на церебральну дисфункцію, але іншим способом (окремо) не локалізується.
Фармакотерапія осцилопсії
Насамперед дуже важливо розуміти різницю між інтрузивними сакадами та ністагмом, щоб призначити оптимальне лікування. Для цього неврологу треба уважно вислухати скарги хворого з нейроофтальмологічними проблемами, з якими він прийшов на консультацію (найчастіша – головокружіння). Так, ністагм – це ініціально аномальний рух, тобто повільний дрейф ока від бажаного положення; він є ритмічним, регулярним, тривалим і стійким. Інтрузивні сакади представлені ініціально нормальним рухом ока, що є неритмічним, нестабільним переміжним, нестійким або викликаним зміною погляду.
Часто пацієнт із нейроофтальмологічними порушеннями скаржиться на осцилопсію – відчуття, що нерухомі об’єкти рухаються в просторі, котре спричиняють ністагми й інтрузивні сакади. Осцилопсія та головокружіння нерідко є супутніми станами. Насамперед потрібно покращити загальний стан хворого, зменшити тривожність. Серед рекомендованих препаратів – анксіолітики: клоназепам, Гамалате В6; габапентин (Ньюропентин); прегабалін (Зонік); баклофен. Анксіолітик позиціонується як один із найкращих засобів у зниженні симптомів тривоги. До того ж такі препарати знижують напруження та мають заспокійливу дію. Що стосується клоназепаму, то його мінус полягає в тому, що ці таблетки потрібно приймати все життя, поступово підвищуючи дозу. Хорошим варіантом заміни є Гамалате В6, що впливає на ГАМК-ергічну систему в цілому та дає можливість зменшити відчуття осцилопсії.
У Гамалате В6 поєднуються чотири природні метаболіти мозку: гамма-аміномасляна кислота (ГАМК), гамма-аміно-бета-оксимасляна кислота (ГАБОМК), магнію глутамат гідробромід (МГГ) та піридоксин (вітамін В6). ГАМК заповнює дефіцит ендогенних ГАМК-ергічних речовин, усуває тривогу й напругу, сприяє позитивній реакції на стресори, покращує концентрацію уваги, пам’ять, когнітивні функції, гальмує ноцицептивну стимуляцію. МГГ блокує глутаматні рецептори, внаслідок чого швидко усуває збудження, тривогу, має релаксувальну дію. Піридоксин бере участь у синтезі ГАМК із глутамінової кислоти, відновлює синтез дофаміну та серотоніну, поліпшує енергообмін нейронів. ГАБОМК – природний метаболіт мозку, що має протисудомний ефект, покращує пам’ять, нормалізує сон, заповнює дефіцит ендогенної ГАМК за її нестачі, виконує функцію холінергічної стимуляції, легше проникає крізь гематоенцефалічний бар’єр.
Анксіолітична дія Гамалате В6 була продемонстрована в дослідженні M. Martinez Mendoza (1992) за участю 100 пацієнтів (середній вік – 41 рік) із симптомами тривоги, в ході якого вивчали наслідки заміни бензодіазепінів (БЗД) на Гамалате В6. Учасників було поділено на дві групи. У групі А хворі не приймали анксіолітичні засоби та мали сильний або тяжкий ступінь тривоги. Протягом 30 днів вони приймали Гамалате В6 по 2 таблетки 3 рази на добу після прийому їжі. У групі Б пацієнти зі значною тривожністю приймали БЗД понад місяць. Потім дозу БЗД знизили в половину й додали ½ дози Гамалате В6. Дію препаратів та ступінь тривожності (визначали за шкалою Гамільтона) оцінювали на початку, на 15-й і 30-й день терапії. Була показана майже вдвічі більша ефективність Гамалате В6 після 30 днів лікування, ніж після 15 днів. При заміні БЗД на Гамалате В6 профіль тривожності не погіршився в жодного пацієнта.
Отже, Гамалате В6 може стати прийнятним замінником БЗД для зниження симптомів тривоги або чергуватися з ними. Заміна БЗД на Гамалате В6 добре переноситься та не зумовлює синдрому відміни. Ефективність препарату аналогічна також для пацієнтів, які приймали чи не приймали анксіолітики.
На завершення своєї доповіді професор М.М. Орос пообіцяв аудиторії, що друга частина лекцій про сакадні системи, зокрема ністагм, буде не менш цікавою та пізнавальною.
Підготувала Олена Коробка