


2 квітня, 2019
Метформін: фармакогенетика, вплив на кишкову мікробіоту та імунітет
В усіх сучасних алгоритмах з лікування цукрового діабету (ЦД) 2 типу препаратом першої лінії є метформін: за відсутності протипоказань він має призначатись усім пацієнтам, які потребують фармакотерапії. Це пояснюється тим, що серед 12 класів засобів для лікування ЦД 2 типу лише метформін поєднує три кардинальні якості, властиві гарним лікам, – надзвичайну безпеку, ефективність та економічну доступність. Дійсно, метформін добре переноситься: лише 5% пацієнтів припиняють його прийом через побічні явища. Загалом метформін є високоефективним у лікуванні ЦД 2 типу й одним з небагатьох пероральних цукрознижувальних препаратів, здатних сприяти зменшенню маси тіла. Крім ЦД 2 типу, метформін використовується для лікування синдрому полікістозних яєчників, і нині є підстави сподіватися, що незабаром його застосування буде розширене на інші показання, як-от профілактика й лікування раку.
Історичні аспекти
Метформін був отриманий з Galega officinalis (козлятник лікарський), традиційної лікарської рослини, що широко застосовувалася у Європі в добу Середньовіччя для полегшення поліурії. Наприкінці XIX ст. було встановлено, що G. officinalis містить велику кількість гуанідину – сполуки, в якої згодом були виявлені гіпоглікемічні властивості. Однак клінічному застосуванню гуанідину перешкоджала його виражена токсичність, тож дослідники перемістили фокус на пошук більш безпечних аналогів. Бігуаніди, що складаються з двох N‑пов’язаних гуанідинів, були синтезовані у 1920-х рр., проте їхній терапевтичний потенціал залишався без належної уваги внаслідок появи в тому ж десятилітті інсулінотерапії. І тільки в 1957 р., після публікації результатів успішного дослідження французького лікаря Жана Стерна, метформін почав використовуватися для лікування діабету. На відміну від раніше застосовуваних протидіабетичних засобів перевагою бігуанідів була здатність знижувати рівні глюкози в крові без ризику розвитку гіпоглікемії. Спочатку популярними були сильніші бігуаніди фенформін і буформін, проте після великої кількості повідомлень про асоційований лактатацидоз ці препарати в 1970-х рр. були відкликані з ринку в більшості країн. У підсумку завдяки гарному профілю безпеки метформін став препаратом першої лінії терапії ЦД 2 типу і сьогодні входить до переліку життєво важливих ліків Всесвітньої організації охорони здоров’я.
Механізм дії та молекулярні мішені метформіну
Механізм дії метформіну відрізняється від такого всіх інших класів пероральних гіпоглікемічних препаратів. Здатність метформіну одночасно пригнічувати глюконеогенез у печінці й інші анаболічні шляхи, зокрема біосинтез ліпідів і холестерину, пов’язана з інгібуванням комплексу I дихального ланцюга мітохондрій і непрямою активацією сигнального шляху енергетично чутливої АМФ-активованої протеїнкінази (AMPK). Головною молекулярною мішенню метформіну вважають мітохондрії, де він пригнічує комплекс I електронно-транспортного ланцюга, що зменшує окисне фосфорилювання і зрештою знижує синтез АТФ.
Підвищення АМФ під дією метформіну інгібує цАМФ-залежну протеїнкіназу А і фруктозо‑1,6-бісфосфатазу; цей механізм забезпечує пригнічення глюконеогенезу. Завдяки активації AMPK клітини переходять з анаболічного стану в катаболічний.
Метформін транскрипційно регулює гени, що кодують глюкозні транспортери в м’язових клітинах (GLUT1); це підвищує захоплення глюкози м’язовими клітинами, знижує рівні глюкози й інсуліну в крові при ЦД 2 типу. Зменшення печінкового глюконеогенезу під дією метформіну непрямим шляхом також знижує циркулюючі рівні інсуліну й інсулін/IGF1-рецепторопосередковану активацію сигнального шляху фосфатидилінозитол‑3-кінази (PI3K).
Протипухлинні ефекти метформіну значною мірою опосередковуються активацією сигнального шляху LKB1/AMPK. LKB1 є пухлинним супресором, і мутація гена LKB1 асоціюється із синдромом Пейтца-Єгерса – вродженим захворюванням зі схильністю до розвитку злоякісних новоутворень. Було продемонстровано, що протекторний ефект метформіну послаблюється при фармакологічному пригніченні AMPK і в експериментальних моделях в AMPK‑нокаутованих тварин. Активація AMPK має наслідком пригнічення сигнального шляху mTOR (мішені рапаміцину ссавців) – нутрієнт-чутливого регулятора синтезу білка, розвитку і проліферації клітин. Цей ефект досягається шляхом прямого фосфорилювання Ser1345 і Thr1227 пухлинного супресора TSC2 (білка туберозного склерозу-2), який формує mTOR‑комплекс-1 (mTORC1) з TSC1. Крім того, AMPK може запобігати активації mTORC1 за допомогою фосфорилювання його сполучного партнера Raptor.
Активація AMPK супроводжується зменшенням ефекту Варбурга – набутого метаболічного фенотипу ракових клітин, який характеризується перевагою аеробного гліколізу над окислювальним фосфорилюванням. Ця трансформація полегшується гіпоксія-індуцибельним фактором 1α – транскрипційним фактором, який активує експресію гліколітичного гена у відповідь на передачу сигналів mTORC1. Класичною метаболічною характеристикою раку, пов’язаною з ефектом Варбурга, є підвищений синтез жирних кислот de novo і підвищені рівні ключового ліпогенного ферменту – синтази жирних кислот (FAS), описані при різних типах раку в людини. Встановлено, що активація AMPK знижує експресію FAS у клітинах раку передміхурової залози і зменшує їх виживання. У дослідженні на моделі карциноми товстої кишки було продемонстровано, що метформін знижує експресію FAS і протидіє стимулюючому впливу висококалорійної їжі на ріст пухлини.
Фармакогенетика метформіну
У пацієнтів із ЦД 2 типу, які отримують ідентичну цукрознижувальну терапію, відрізняються показники глікемічного контролю, частота побічних ефектів і комплаєнс. Це значною мірою пов’язано з поліморфізмом генів, що беруть участь в абсорбції, транспортуванні та метаболізмі пероральних гіпоглікемічних засобів.
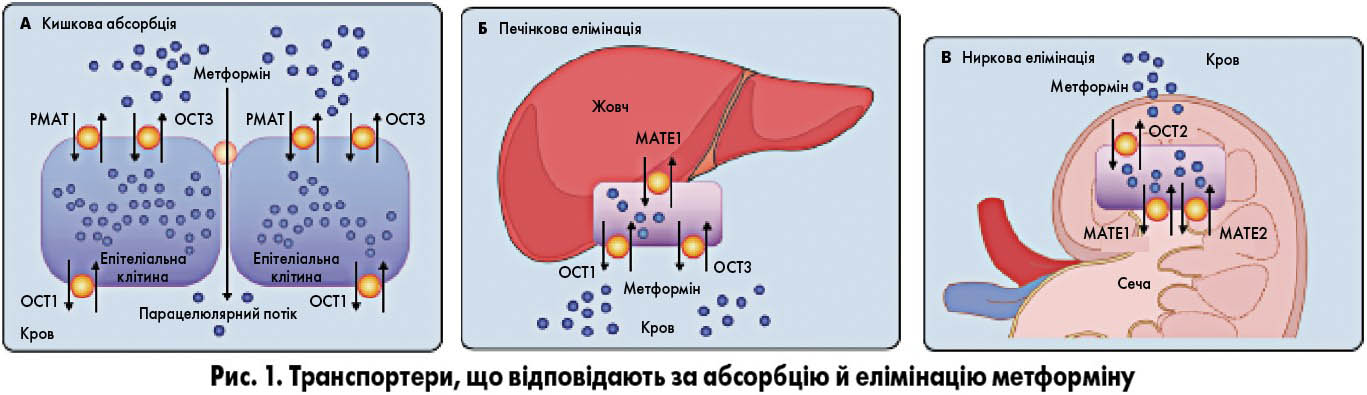
Незвична гідрофільна природа метформіну унеможливлює його пасивний транспорт крізь клітинні мембрани. В ентероцити, гепатоцити й ниркові епітеліальні клітини метформін активно транспортується органічними катіонними транспортерами (OCT).
Після перорального прийому метформін досягає кишкового ендотелію (рис. 1, А) і поглинається ентероцитами за допомогою плазматичного моноамінного транспортера (PMAT; кодується геном SLC2A4) та OCT3 (кодується геном SLC22A3), розташованих на люмінальній поверхні клітин. Крізь базолатеральну мембрану ентероцита за допомогою OCT1 (ген SLC22A1) метформін надходить у кровоток. Після потрапляння метформіну в печінку (рис. 1, Б) OCT1 та ОСТ3 забезпечують його поглинання гепатоцитами. Екскреція метформіну в жовч відбувається за участю MATE1 (multidrug and toxin extrusion protein 1, ген SLC47A1). Епітеліальні клітини нирок (рис. 1, В) захоплюють метформін з крові за допомогою OCT2, після чого він за участю МАТЕ1 та МАТЕ2 в незміненому вигляді екскретується із сечею. Наявність генетичних варіантів генів, що кодують ці транспортери, може пояснювати індивідуальні особливості фармакогенетики і фармакодинаміки метформіну.
З метою вивчення наявності асоціації різних варіантів гена ОСТ SLC22A1, SLC22A та SLC47A1 з цукрознижувальним ефектом метформіну було проведено клінічне дослідження за участю пацієнтів із ЦД 2 типу, які раніше не отримували метформін. Результати показали тенденцію до більшого зниження рівня глікованого гемоглобіну (HbA1c) в пацієнтів, гомозиготних за А‑алеллю гена SLC47A1 rs2289669, порівняно з носіями G‑алелі цього гена (p=0,059).
У дослідженні, проведеному в Нідерландах, вивчали взаємозв’язок між однонуклеотидними поліморфізмами (SNP) гена SLC22A1 і цукрознижувальним ефектом метформіну в осіб європейської популяції. У дослідження було включено 102 пацієнти віком понад 55 років з великого проспективного когортного дослідження Rotterdam Study, у яких визначали рівень HbA1c до початку лікування і потім після 30 і 120 днів терапії. Було встановлено, що SNP у точці rs622342 гена SLC22A1 асоціюється із цукрознижувальним ефектом метформіну, а саме в носіїв генотипу CC відзначалося на 58% більш виражене зниження HbA1c, ніж у носіїв генотипу AA. При цьому частка хворих, які припинили лікування метформіном і розпочали прийом інших препаратів, була більшою серед носіїв генотипу АА.
Вивчення інтронного варіанта гена rs2289669 (G>A) гена SLC47A1, що кодує MATE1, продемонструвало більш виражене зниження рівня HbA1c у пацієнтів, які використовували метформін. Водночас у дослідженні DPP показано, що наявність варіанта rs8065082 (C>T) гена SLC47A1 асоціювалася з вищими шансами профілактики діабету в осіб, які отримували метформін.
За допомогою технології повногеномних асоціативних досліджень (Genom Wide Association Studies – GWAS) із застосуванням даних пацієнтів двох великих досліджень – UKPDS і GoDARTS – було ідентифіковано SNP rs11212617 поруч з геном атаксії-телеангіектазії (ATM), асоційованим з ефективністю лікування метформіном. Ген АТМ бере участь у процесах репарації ДНК і контролю клітинного циклу, а також відіграє роль у впливі метформіну на AMPK; зміни в цьому гені, ймовірно, впливають на тривалість гіпоглікемічного ефекту метформіну.
У метааналізі 10 557 пацієнтів, проведеному Консорціумом MetGen, за допомогою GWAS було встановлено статистично значимий зв’язок SNP rs8192675 (інтрона глюкозного транспортера GLUT2, який кодується геном SLC2A2 й експресується в гепатоцитах) з рівнем HbA1c на тлі лікування метформіном (абсолютна різниця між двома гомозиготними групами – 0,33%). Цікаво, що алель, асоційована з кращою відповіддю на метформін, також корелює з більш високим вихідним HbA1c. Загалом комбінований вплив локусів ATM та SLC2A2 на цукрознижувальну ефективність метформіну є мінімальним, що свідчить про необхідність продовження пошуку генетичних детермінантів відповіді на цей препарат.
Метформін і кишкова мікробіота
Зміни кишкової мікробіоти, спричинені дієтою, антибіотиками й іншими чинниками, пов’язані з розвитком багатьох захворювань, як-от ожиріння, діабет, метаболічний синдром і рак, а також із прискореним старінням. Є докази того, що метформін може модулювати склад і метаболізм кишкової мікробіоти, наприклад зменшувати кількість Instinibacter та підвищувати продукцію бутирату. Питання про те, якою мірою зміни мікробіоти пояснюють сприятливі ефекти метформіну при метаболічних захворюваннях, продовжує вивчатися.
Однією з моделей для дослідження взаємозв’язку макроорганізму з кишковою мікробіотою є Caenorhabditis elegans – нематода, що сформувала вигідні взаємини з E. coli. Було встановлено, що в С. elegans метформін змінює бактеріальний метаболізм фолату і метіоніну, імітуючи дію обмеження енергетичної цінності їжі. При цьому метформін уповільнював процес старіння тільки в присутності E. coli.
ЦД 2 типу й ожиріння характеризуються структурними та функціональними змінами популяцій кишкових мікробів, запальним станом і порушенням бар’єрної функції кишечнику. Схожість ефектів мікробіоти і метформіну на ожиріння та регуляцію глюкози крові вказує на існування загальних механізмів, при цьому протидіабетичні ефекти метформіну досягаються як прямим шляхом, так й опосередковано завдяки впливу на кишкову мікробіоту.
У двох експериментальних дослідженнях було встановлено, що в умовах дієти з високим умістом жирів метформін покращує метаболічні маркери і наближає склад фекальної мікробіоти до такого в умовах нормального харчування. За допомогою метагеномного аналізу було продемонстровано, що метформін індукує глибокі зрушення специфічних підгруп бактерій, зокрема збільшує вміст мікроорганізмів роду Akkermansia. Крім того, метформін підвищує кількість келихоподібних клітин, що продукують муцин, який є джерелом харчування для Akkermansia municiphila. Відомо, що призначення цієї бактерії як пробіотичного препарату асоціюється з поліпшенням метаболічного профілю, зменшенням метаболічної ендотоксемії і запалення жирової тканини, покращенням глікемічного контролю і зниженням інсулінорезистентності; всі ці ефекти також властиві метформіну.
На тлі дієти з високим умістом жирів метформін збільшує популяцію лактобацил. Lactobacillus є однією з багатьох бактерій, які можуть утилізувати глюкозу з утворенням лактату. Слід зауважити, що покращення гомеостазу глюкози при призначенні метформіну істотно послаблюється на тлі супутнього застосування антибіотиків широкого спектра дії. Ці дані свідчать про те, що кишкова мікробіота є терапевтичною мішенню для метформіну. Імовірно, підвищені рівні лактату й інших субстратів для глюконеогенезу (пірувату, аланіну) при лікуванні метформіном не тільки захищають макроорганізм від гіпоглікемії, а й модифікують структуру мікробіоти, надаючи нові енергетичні субстрати певним бактеріям.
Комбінована терапія метформіном і пребіотічними харчовими волокнами може покращувати метаболічний профіль, особливо в разі дієти з високим умістом жирів. При цьому пребіотики й метформін виявляли синергічну й адитивну сприятливу дію на масу тіла, печінкові тригліцериди, секрецію глюкозозалежного інсулінотропного поліпептиду й лептину, а також на активацію AMPK; водночас значно збільшувався вміст біфідогенних бактерій, зокрема Bifidobacterium spp.
Метформін також може впливати на системний метаболізм вуглеводів за допомогою гастроінтестинальних механізмів, не пов’язаних безпосередньо з кишковою мікробіотою. Ці механізми включають модулювання фізіології жовчних кислот, кишкових гормонів і дуоденальної AMPK, що бере участь у пригніченні глюконеогенезу в печінці.
Метформін та імунна система
Клінічний досвід застосування метформіну в пацієнтів із ЦД 2 типу не надав доказів впливу прийому цього препарату на імунну функцію, оскільки ні імунодефіцит, ні підвищення аутоімунітету не реєструвалися як побічні реакції. Проте дані молекулярних досліджень, а також здатність позитивно впливати на розвиток і перебіг захворювань, не пов’язаних з порушенням вуглеводного обміну, свідчать про наявність у метформіну імуномодулювальних властивостей.
На рисунку 2 схематично зображений вплив метформіну на імунні клітини та пов’язані з ними фібробласти. Після потрапляння в клітину метформін реалізує низку AMKP‑залежних і AMPK‑незалежних ефектів, зокрема зменшує утворення активних форм кисню (АФК) та нейтрофільних екстрацелюлярних пасток (NET); пригнічує сигнальні шляхи mTOR/STAT‑3 та TGFβ/Smad.
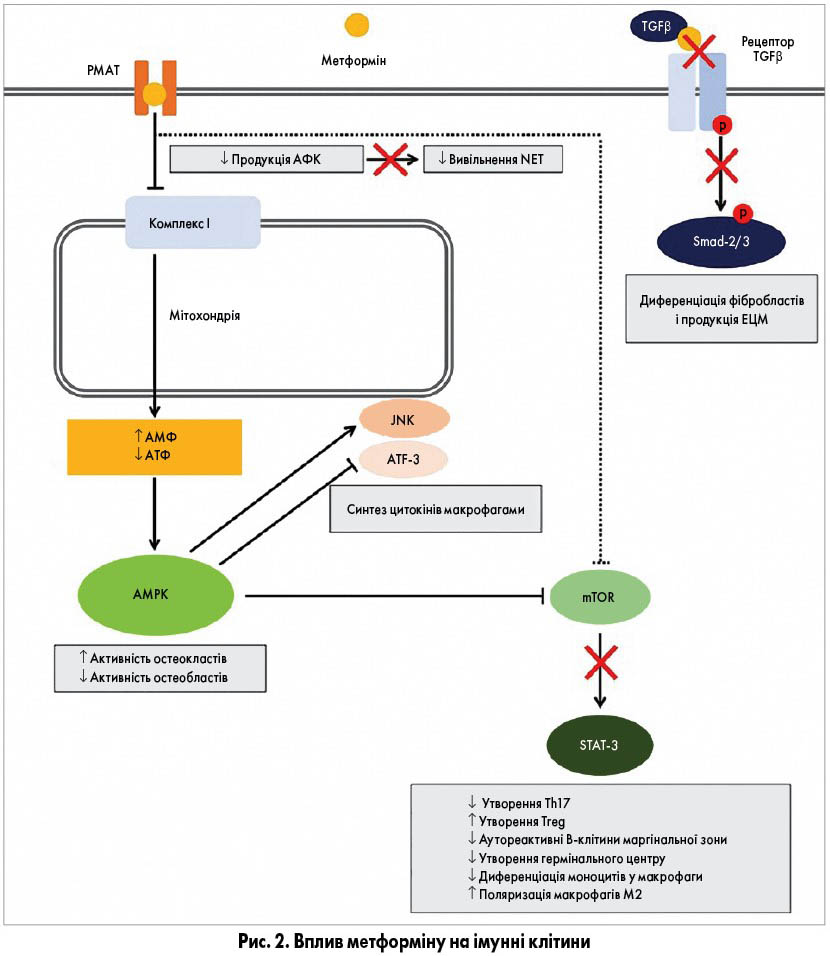
В експериментальних дослідженнях метформіну як антинеопластичного агента в інтактних та імуноскомпрометованих мишей протипухлинна активність препарату була значно вищою у тварин з функціонуючою імунною системою, в яких метформін AMPK‑залежно збільшував кількість CD8+ лімфоцитів, що інфільтрують пухлину. В інших дослідженнях було встановлено, що метформін може посилювати активність імунотерапевтичних протипухлинних препаратів – блокаторів білка програмованої клітинної смерті-1 (PD‑1), імовірно, шляхом зменшення споживання кисню внаслідок пригнічення окисного фосфорилювання. Крім того, вплив метформіну на імунологічну пам’ять асоціювався з підвищенням ефективності протипухлинної вакцини.
Метформін дозозазалежно інгібує вивільнення IL‑8, індуковане IL‑1β, в непосмугованих м’язових клітинах судин, макрофагах й ендотеліоцитах. Цей прямий протизапальний вплив на судинну стінку забезпечується шляхом зниження транслокації ядерного фактора транскрипції каппа B (NF‑κB).
Під час вивчення впливу метформіну на низку показників запалення й імунітету в пацієнтів із ЦД 2 типу та кардіоваскулярними захворюваннями було встановлено, що монотерапія метформіном тривалістю від 8 до 12 міс на відміну від лікування похідними сульфонілсечовини сприяла зниженню інтенсивності системного низькорівневого запального процесу, що підтверджувалося достовірним зниженням співвідношення нейтрофілів до лімфоцитів у периферичній крові. Одночасно в групі метформіну спостерігалося значне зниження рівня прозапальних цитокінів (IL‑1β, IL‑6, хемокінів CXCL1/2) у плазмі крові, що супроводжувалося покращенням клінічного стану пацієнтів.
У пацієнтів із розсіяним склерозом метформін зменшує запалення в центральній нервовій системі та може покращувати перебіг захворювання. Експериментальні дані свідчать, що метформін модулює метаболізм CD4+ Т‑лімфоцитів і таким чином зменшує аутоімунну відповідь.
Висновки
Незважаючи на те, що протягом останніх років арсенал лікарських засобів для лікування ЦД 2 типу поповнився великою кількістю нових засобів, метформін залишається препаратом першої лінії згідно з усіма світовими рекомендаціями. Це пояснюється його ефективністю, безпекою й економічною доступністю.
У низці досліджень було продемонстровано, що на тлі лікування метформіном відбуваються кількісні та якісні зміни кишкової мікробіоти, які сприяють протидіабетичним ефектам препарату, особливо в пацієнтів з надлишковою масою тіла чи ожирінням.
 Переконливі експериментальні дані свідчать про наявність у метформіну імуномодулювальних властивостей, що можуть бути корисними в профілактиці й лікуванні злоякісних новоутворень, аутоімунних, кардіоваскулярних і нейродегенеративних захворювань.
Переконливі експериментальні дані свідчать про наявність у метформіну імуномодулювальних властивостей, що можуть бути корисними в профілактиці й лікуванні злоякісних новоутворень, аутоімунних, кардіоваскулярних і нейродегенеративних захворювань.
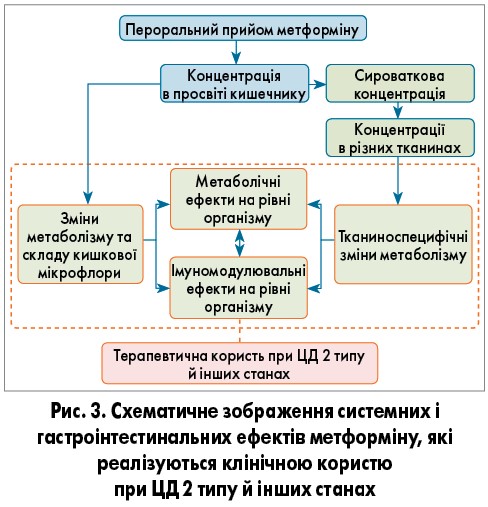
Глікемічні та неглікемічні механізми дії метформіну, які реалізуються в кишечнику, інших тканинах, забезпечують метаболічні й імуномодулювальні ефекти на рівні всього організму, що пояснює клінічну користь від застосування препарату при діабеті та інших патологічних станах (рис. 3).
Вивчення поліморфізму генів, що відповідають за дію метформіну в пацієнтів із ЦД 2 типу, є важливим інструментом персоналізації ефективної та безпечної фармакотерапії.
Список літератури знаходиться в редакції.
Підготував Олексій Терещенко
Медична газета «Здоров’я України 21 сторіччя» № 5 (450), березень 2019 р.