


30 жовтня, 2020
Перспективи використання ксиліту як засобу для селективного пригнічення проліферації ракових клітин
Пошук продуктів або добавок із протираковими властивостями є перспективним напрямом в онкології, оскільки їх споживання може допомогти запобігти розвитку онкопатології і сприяти опірності організму навіть за наявності злоякісної пухлини. Біологічно активні сполуки рослинного походження все частіше визнаються дієвими компонентами лікування раку. Підбір дієти для онкологічних пацієнтів досі залишається невирішеною проблемою. Тому не випадковий інтерес до вивчення цукрозамінника ксиліту, що є безпечним компонентом харчових продуктів і при цьому виявляє здатність пригнічувати проліферацію ракових клітин. Пропонуємо огляд досліджень, присвячених оцінці онкосупресорних властивостей ксиліту.
N. Tomonobu та співавт. (2020) відзначають, що пошуки нових ефективних протипухлинних препаратів серед традиційних для Азії лікарських засобів привернули увагу до паразитичного гриба Cordyceps militaryaris, відомого своїми протипухлинними властивостями, що зумовлені вмістом природного нуклеозидного антибіотика кордицепіну [29-31]. В екстракті C. militaryaris було також виявлено високий вміст ксиліту, який несподівано для дослідників проявляв протиракову активність [30]. Була висунута гіпотеза про те, що ксиліт може стати корисним компонентом профілактики онкопатології, а також зниження агресивності злоякісної пухлини. Оскільки вартість ксиліту невисока (на відміну від кордицепіну), він легкодоступний і повсюдно використовується в харчових продуктах, що свідчить про високий рівень його безпеки. Однак ще належить з’ясувати конкретний механізм, за допомогою якого ксиліт викликає загибель ракових клітин.
N. Tomonobu та співавт. (2020) оцінили залежність індукції апоптозу клітин меланоми від дози ксиліту, починаючи з 2 мг/мл як найнижчої концентрації. Було підтверджено, що 24-годинна обробка ксилітом викликала дозозалежний апоптоз клітин MeWo (клітинна культура меланоми людини). Доза 50 мг/мл забезпечувала найвищий ефект.
Враховуючи те, що глюкоза в концентрації 50 мг/мл не могла викликати апоптоз у клітинах MeWo через вищий осмотичний тиск (329 мОсм/л) порівняно з фізіологічним (285±5 мОсм/л), N. Tomonobu та співавт. вирішили використовувати концентрацію ксиліту 25 мг/мл як найвищу в серії експериментів in vitro. Для отримання додаткової інформації про клітинні події, що лежать в основі ксиліт-індукованого апоптозу, був досліджений окисний статус клітини.
Як і очікувалося, апоптоз, викликаний ксилітом у концентрації 25 мг/мл, супроводжувався помітним підвищенням вмісту активних форм кисню (АФК). Паралельно було виявлено значне зниження рівня відновленого глутатіону і лише невелике зниження вмісту АТФ. Примітно, що ксиліт-опосередкована апоптотична загибель клітин була значно ослаблена попередньою обробкою інгібітором АФК N-ацетил-L-цистеїном; це дозволяє припустити, що АФК активно залучені до ксиліт-опосередкованого апоптотичного процесу ракових клітин.
За допомогою кількісної полімеразної ланцюгової реакції у режимі реального часу встановлено, що ксиліт порівняно з кордицепіном набагато сильніше стимулює ген CHAC1, який може брати участь у регуляції апоптозу.
Роль гена CHAC1 у ксиліт-опосередкованій загибелі ракових клітин
У роботі N. Tomonobu та співавт. (2020) вперше було показано, що індукція ферменту CHAC1 (ChaC – глутатіонспецифічна γ-глутамілциклотрансфераза 1) викликає деградацію глутатіону і є основною причиною ініційованої ксилітом апоптотичної загибелі ракових клітин.
Щоб перевірити, чи обмежений ксиліт-опосередкований апоптоз злоякісними клітинами MeWo (і чи не впливає ксиліт на нормальні клітини), N. Tomonobu та співавт. досліджували вплив ксиліту на інші типи ракових клітин людини і нормальні людські фібробласти. Було виявлено, що клітини раку підшлункової залози людини (PANC‑1) демонстрували апоптотичний фенотип, подібний до такого у клітин MeWo після обробки ксилітом, тоді як фібробласти (OUMS‑24) не мали будь-якого помітного апоптотичного фенотипу. Селективний апоптоз ракових клітин був додатково підтверджений аналогічними незалежними експериментами на інших лініях клітин раку підшлункової залози (AsPC‑1 і MIA PaCa‑2). Крім того, в лініях ракових клітин постійно продукувалися АФК на значному рівні, проте в нормальних клітинах OUMS‑24 цієї індукції не відбувалося, і серед них не було також виявлено індукції апоптотичних клітин. Частота опосередкованих ксилітом апоптотичних подій узгоджувалася з частотою індукції CHAC1 у всіх досліджених типах клітин (OUMS‑24, MeWo і PANC‑1).
Потім шляхом пригнічення експресії гена CHAC1 оцінювали його значущість для апоптотичної загибелі клітин. Результати показали, що зниження активності гена CHAC1 призводило до значного ослаблення зумовленої ксилітом апоптотичної загибелі клітин, пригнічення ксиліт-опосередкованої продукції АФК і відновлення рівня глутатіону. Таким чином, з урахуванням того, що CHAC1 є основним ферментом, який руйнує глутатіон, ксиліт-опосередковане збільшення рівня АФК видається раціональним (Crawford et al., 2015).
Ксиліт індукує стрес ендоплазматичного ретикулуму
Повідомлялося, що індукція CHAC1 транскрипційно регулюється за допомогою активуючого фактора транскрипції 4 (АТФ4) через стресовий вплив на ендоплазматичний ретикулум (ЕПР), і тому була досліджена потенційна здатність ксиліту викликати стрес ЕПР [33-35]. Індукція основного ЕПР-шаперона – білка, який зв’язує імуноглобулін (BiP), є надійним індексним маркером для спостереження клітинних подій після стресу ЕПР. За відсутності стресу ЕПР у клітинах BiP зв’язує і пригнічує три репрезентативних стрес-сенсори ЕПР: інозитолзалежний білок 1α (IRE1α), який активує фактор транскрипції 6 і PKR-подібну протеїнкіназу ЕПР. В умовах ЕПР-стресу BiP рекрутується неправильно згорнутими білками і згодом звільняється від трьох сенсорів [16]. У результаті вільні сенсори зазнають автоактивації та індукують власні сигнальні каскади, причетні до множинних клітинних подій стресу ЕПР. N. Tomonobu та співавт. спробували визначити, чи реагують клітини на стрес ЕПР, оцінюючи рівні індукції BiP після обробки клітин ксилітом.
Було виявлено, що ксиліт, але не глюкоза, значущо індукував білок BiP паралельно з індукцією CHAC1 залежно від дози. Крім того, при використанні АТФ4-специфічного зонда для оцінки стану активації АТФ4 після обробки ксилітом було показано, що саме АТФ4, але не ядерний фактор κB значною мірою активується у відповідь на вплив ксиліту, причому цей процес подібний до індукції білків BiP і CHAC1.
Таким чином, індукція CHAC1 є необхідним стресорним фактором для ЕПР, тригером якого в ракових клітинах є ксиліт і який пов’язаний із вторинною індукцією оксидативного стресу в оброблених клітинах, що зрештою призводить до апоптотичної загибелі клітин.
Ксиліт зменшує ріст меланоми in vivo через стрес ЕПР
Оскільки відомо, що ксиліт сприяє селективній загибелі ракових клітин in vitro, N. Tomonobu та співавт. досліджували, чи діє ксиліт подібним чином in vivo. В експериментальних умовах виявилося, що при пероральному введенні ксиліт не впливав на об’єм і масу пухлини, яка походить із клітин MeWo, тоді як внутрішньовенна ін’єкція послаблювала швидкість росту пухлини (спостерігалося зменшення її об’єму та маси – рис.). Мишам вводили ксиліт у дозі 2,0 г/кг, оскільки вона аналогічна ефективній і максимальній дозі in vitro – 25 мг/мл.
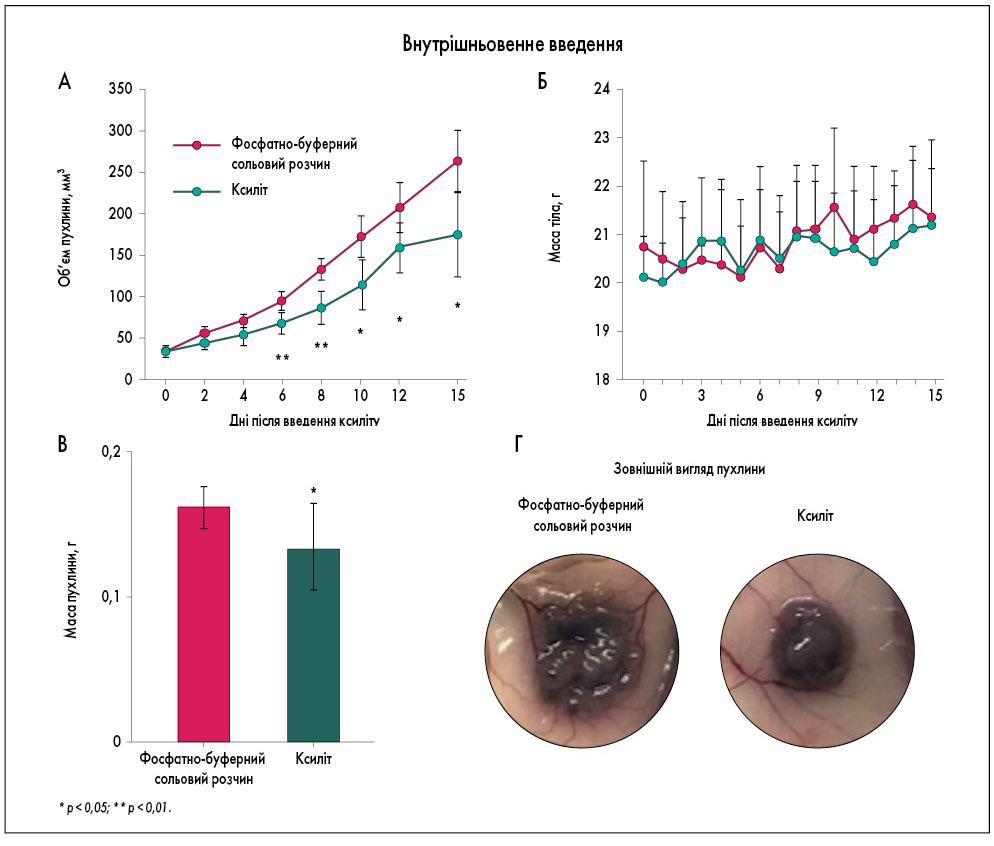 Рис. Протипухлинна дія ксиліту in vivo
Рис. Протипухлинна дія ксиліту in vivo
Адаптовано за Tomonobu N. Xylitol acts as an anticancer monosaccharide to induce selective cancer death via regulation of the glutathione level. Chem Biol Interact. 2020 Jun 1; 324: 109085.
При дослідженні індукції стресу ЕПР і подальшого статусу CHAC1 in vivo виявлялося значне накопичення BiP, а також CHAC1 у пухлинах саме при внутрішньовенному, але не пероральному введенні ксиліту.
Вирішальну роль у пригніченні пухлини, викликаному обробкою ксилітом, в умовах як in vivo, так і in vitro, ймовірно, відіграє стрес ЕПР. Це було підтверджено використанням основного інгібітора ЕПР-стресу – тауроурсодеоксихолевої кислоти (TUDCA), яка ефективно «скасовувала» ксиліт-опосередковане пригнічення пухлини: на 15-й день маса пухлин, оброблених ксилітом, досягала майже тих же рівнів, що і в контрольних групах (обробка фосфатно-буферним сольовим розчином).
Ці результати показують, що в разі внутрішньовенного введення ксиліту притаманна функція пригнічення пухлини, і ця функція залежить від наявності певного рівня стресу ЕПР.
Ксиліт сенсибілує клітини меланоми до хіміотерапевтичних препаратів
Оксидативний стрес, викликаний ксиліт-опосередкованим відновленням глутатіону, може посилювати дію хіміотерапевтичних препаратів. Раніше було показано, що ксиліт помітно підсилює опосередкований кордицепіном апоптоз ракових клітин [30]. У даному дослідженні продовжили вивчати цю дію. Як і очікувалося, ксиліт ефективно сенсибілізував клітини меланоми до протиракової сполуки 5‑флуороурацилу (5‑FU) як in vitro, так і in vivo. Спостерігався також ефект сенсибілізації ксилітом in vitro для іншої сполуки, дакарбазину, клінічно схваленого для лікування меланоми. Таким чином, результати показали, що ксиліт підсилює дію хіміотерапевтичних препаратів.
Обговорення й висновки
Результати дослідження показали, що ксиліт має помітну здатність пригнічувати ріст ракових клітин як при культивуванні in vitro, так і на мишачій моделі меланоми in vivo. Пероральне введення ксиліту тваринам не сприяло онкосупресорній дії (ймовірно, через кислотно-опосередковану модифікацію/деградацію ксиліту або його недостатнє надходження у кров з травного каналу), тоді як його внутрішньовенне введення запобігало росту пухлини. Ксиліт-опосередкований апоптоз ракових клітин був викликаний активацією CHAC1 (чому також сприяв викликаний ксилітом стрес ЕПР) і оксидативним стресом, оскільки CHAC1 ферментативно розщеплював внутрішньоклітинний глутатіон.
Ці процеси спостерігалися в основному для клітин MeWo; деякі ключові результати були підтверджені для інших типів ракових клітин (наприклад, PANC‑1, AsPC‑1 і MIA PaCa‑2). Таким чином, запропонований апоптотичний шлях, опосередкований ксилітом, не обмежений тільки однією клітинною лінією, проте, судячи з усього, не діє в нормальних фібробластах OUMS‑24 людини. Це дає підстави вважати, що ксиліт може бути ефективним і корисним щоденним харчовим компонентом для селективного запобігання розвитку раку.
Пригнічення експресії CHAC1 викликало помірне зниження зумовленої ксилітом продукції АФК у клітинах MeWo. Цей результат може вказувати на те, що активація CHAC1 стресом ЕПР не є єдиним джерелом АФК у клітинах, оброблених ксилітом. Аналогічно, активація CHAC1 може бути не єдиною подією, відповідальною за протиракові властивості ксиліту, як показано в дослідженнях in vivo.
Сьогодні не відомо, як ксиліт викликає ЕПР-стрес у ракових клітинах. Дати підказку може вивчення внутрішньоклітинних метаболітів ксиліту. Повідомлялося, що один із них, ксилулоза‑5‑Р, здатний пригнічувати систему гліколізу в клітинах ссавців [36]. Таким чином, рівень АТФ, що виробляється при гліколізі і необхідний для згортання білка, може бути значно знижений шляхом обробки ксилітом. Однак результати дослідження показали, що дія ксиліту не викликала споживання АТФ на значно вищому рівні в оброблених ним клітинах. У додаткових дослідженнях на апоптотичну загибель клітин при дії ксиліту не впливало гіпоксичне середовище, яке зміщує синтез АТФ у мітохондріях у бік його продукції на основі гліколізу.
Було висунуто припущення, що ксиліт може перешкоджати нормальному глікозилюванню білка в ЕПР. Оскільки ксиліт має рослинне походження, він може конкурувати з глюкозою або іншим моносахаридом, який використовується для реакцій глікозилювання білків у ЕПР. Раніше було показано, що профіль глікозилювання змінюється при обробці ксилітом і це може супроводжуватися помітним збільшенням неправильно згорнутих білків, які пов’язані зі стресом ЕПР. Цю ідею може підтвердити відкриття того факту, що основний індуктор ЕПР‑стресу, тунікаміцин, опосередковував неповне глікозилювання білків, яке може бути причиною їх неправильного згортання [37].
Ще одне питання полягає в тому, чим зумовлена селективність щодо ракових клітин при стресі, пов’язаному з ксилітом.
N. Tomonobu та співавт. вважають, що не слід ігнорувати можливе пояснення різних реакцій на ксиліт у нормальних і пухлинних клітинах, наприклад, відмінності у внутрішньоклітинній «поведінці» ксиліту та його метаболітів, оскільки низка ферментів, задіяних у перетворенні ксиліту, неминуче наявна при різних профілях експресії й активності в різних типах клітин. Молекули, які мають стійкі функції щодо ЕПР-стресу або апоптозу, також можуть робити свій внесок. Узагальнені результати підкреслюють необхідність подальшого всебічного аналізу мембранних транспортерів, метаболітів ксиліту і внутрішньоклітинних білків для розширення розуміння складних механізмів чутливості та стійкості до ксиліту, що своєю чергою може пояснити, чим зумовлена селективність або специфічність клітин щодо ксиліту.
У дослідженні Y.W. Naguib та співавт. (2014) отримано дані, які показують, що протипухлинний ефект ксиліту є помірним, але ксиліт має певну здатність посилювати дію часто застосовуваних протипухлинних препаратів, таких як 5-FU.
Підбиваючи підсумки, автори зазначають, що отримані результати підтверджують думку про онкосупресорну дію ксиліту через оксидативний стрес. Ксиліт викликає ЕПР‑стрес, що призводить до індукції CHAC1 і зниження вмісту глутатіону в клітинах. Опосередкований ксилітом оксидативний стрес значною мірою сприяє посиленню протипухлинного ефекту хіміотерапевтичного препарату 5‑FU як in vitro, так і in vivo. Дослідники дійшли висновку, що хіміотерапія в поєднанні з ксилітом може ефективно поліпшувати результати лікування пацієнтів з онкопатологією.
Довідка ЗУ
Препарати для інфузії, виготовлені на основі розчинів ксиліту, схвалені для застосування в Україні та вже використовуються в онкології й інтенсивній терапії [1-4].
Комплексний інфузійний препарат Ксилат® – багатокомпонентний поліфункціональний збалансований розчин. Склад: 100 мл розчину містить 5,0 г ксилітолу, 0,26 г натрію ацетату, 0,6 г натрію хлориду, 0,01 г кальцію хлориду, 0,03 г калію хлориду, 0,01 г магнію хлориду. При внутрішньовенному введенні Ксилат® швидко включається в загальний метаболізм, 80% засвоюється у печінці і накопичується у вигляді глікогену, стимулює окиснення жирних кислот некетогенним шляхом, сприяє утилізації кетонових тіл у циклі Кребса. Препарат не спричиняє зниження в печінці рівня нуклеотидів (АТФ, АДФ, АМФ), безпечний для введення хворим із дефіцитом ферменту фруктозо-1,6-дифосфатази або непереносимістю фруктози. Завдяки складу Ксилат® чинить енергетичну, антикетогенну, анаболічну, антиоксидантну, дезiнтоксикацiйну, гемодинамічну, реологiчну, олужнювальну дію.
Препарат широко застосовується в інфузійній терапії для зменшення явищ інтоксикації, поліпшення мікроциркуляції, корекції кислотно-основного стану, поліпшення гемодинаміки при травматичному, операційному, гемолітичному й опіковому шоку, гострій крововтраті, у процесі передопераційної підготовки і в післяопераційний період, при некомпенсованому цукровому діабеті й кетоацидозі. Обмеженнями в застосуванні розчину Ксилат® є гіперосмолярна кома, анурія, а також клінічні ситуації, коли протипоказане введення великої кількості рідини.
Однією з основних діючих речовин розчину Ксилат® є ксилітол (ксиліт) у концентрації 50 мг/мл, що відкриває потенційно нові можливості застосування препарату у широкого кола онкологічних пацієнтів.
- Ксилат® розчин – інструкція. https://compendium.com.ua/info/168255/ksilat-sup-sup-/.
- Галушко О.А., Шлапак І.П. Новий комплексний інфузійний препарат ксилат і показання до його клінічного застосування (Інформаційний лист). https://www.uf.ua/ua/endokrynolog/vozmozhnostyksylata-v-lechenyy-atsetonemyches-kogo-syndroma-u-detej.
- Курсов С.В., Никонов В.В. Стрессовая гипергликемия: обсуждение пути ее устранения с помощью применения многоатомных спиртов // Медицина неотложных состояний. – 2019. – № 4 (99). – С. 30-37.
- Грівкова Л.В. Патофізіологічне обґрунтування і перспективи використання ксиліту в нутритивній підтримці онкологічних пацієнтів // Практична онкологія. – 2019. – Т. 2, № 4.
Матеріал підготовлений за підтримки фармацевтичної компанії «Юрія-Фарм».
Список літератури знаходиться в редакції.
Тематичний номер «Онкологія, Гематологія, Хіміотерапія» № 4 (65) 2020 р.
