


22 липня, 2016
Гипоталамо-пролактиновая ось. Часть 1: история изучения, нейроэндокринный контроль секреции пролактина
К 60-летию нейроэндокринологии
В 1948 г., когда Джефри Харрис опубликовал свою фундаментальную монографию «Невральный контроль гипофиза», уже было известно, что пролактин, или «лактогенный гормон», как он тогда назывался, может контролироваться рядом других гормонов гипофиза. Харрис разработал убедительную концепцию, согласно которой гуморальные факторы гипоталамуса контролируют секрецию гормонов передней доли гипофиза, правильно предсказав существование гипоталамических рилизинг-факторов, опосредующих невральный контроль секреции гипофиза (Harris, 1948).
Несмотря на наличие очевидных доказательств того, что невральные стимулы (например, кормление грудью) могут стимулировать секрецию пролактина, Харрис считал, что пересечение воронки гипоталамуса не прерывает лактацию (Dempsey & Uotila, 1940) и, следовательно, гипоталамический рилизинг-фактор не является необходимым для стимуляции секреции пролактина. Впоследствии было установлено, что у мышей с удаленным гипофизом эктопические гипофизарные трансплантаты могут поддерживать функцию желтого тела (Everett, 1954) и лактацию (Cowie et al., 1960), т.е. была продемонстрирована гипоталамо-независимая секреция пролактина трансплантатами. На первый взгляд, это противоречит концепции неврального контроля передней доли гипофиза, предложенной Харрисом. Однако дальнейшие исследования показали, что гипоталамическая регуляция действительно необходима для нормальной секреции пролактина (как и предсказывал Харрис), но механизм этого контроля иной.
Эверетт в экспериментах на грызунах обнаружил, что гипофизарная ткань, трансплантированная в отдаленные от гипоталамуса области, индуцирует псевдобеременность (маркер повышенной секреции пролактина), однако если трансплантаты гипофиза реваскуляризируются портальными сосудами, находящимися под контролем срединной возвышенности гипоталамуса, нормальный цикл и, следовательно, нормальные низкие уровни пролактина у животных восстанавливаются (Nikitovitch-Winer & Everett, 1958). Две группы исследователей независимо друг от друга продемонстрировали, что экстракты срединной возвышенности или инфундибулярной области гипоталамуса могут ингибировать секрецию пролактина клетками передней доли гипофиза in vitro (Pasteels, 1963; Talwalker et al., 1963).

Позднее было установлено, что поражение срединной возвышенности (Arimura et al., 1972) или разрушение воронки гипоталамуса (Kanematsu & Sawyer, 1973) вызывают повышение секреции пролактина (что является причиной псевдобеременностей, описанных Эвереттом). Таким образом, было доказано, что гипоталамус необходим для регуляции секреции пролактина, однако он обеспечивает преимущественно ингибиторный эффект. В настоящем кратком обзоре обсуждаются современные представления о гипоталамическом контроле секреции пролактина и нейроэндокринных функциях этого гормона.
С самого начала изучения было очевидно, что невральный контроль секреции пролактина и всей гипоталамо-пролактиновой оси отличается от других гипоталамо-гипофизарных систем. В частности, гипоталамическая регуляция секреции пролактина является не стимулирующей, а ингибирующей, и в ней принимает участие катехоламинергический нейротрансмиттер дофамин, а не более типичные пептидные гормоны гипоталамуса, участвующие в регуляции всех остальных гипофизарных систем.
Кроме того, пролактин - единственный гормон передней доли гипофиза, не имеющий эндокринной целевой ткани. Вследствие этого пролактин регулируется не по системе обратной связи, классической для большинства других гормонов, а посредством обратной связи по типу короткой петли, когда пролактин сам стимулирует секрецию ингибирующего фактора (дофамина). Наконец, несмотря на простое и однобокое название, функции пролактина далеко не ограничиваются индукцией и поддержанием лактации. Сегодня пролактин рассматривается как плейотропный гормон с самым широким спектром физиологических эффектов, превышающим таковой любых других сигнальных молекул организма.
| Таблица. Роль пролактина в адаптации материнского организма к беременности | |||
| Ткань или функция | Адаптивные изменения во время беременности | Доказательства потенциальной роли пролактина | Избранные публикации |
| Молочная железа / лактация | Разветвление и альвеолярное развитие молочной железы Секреция молока | Неоднозначная роль (у Prlr -/- мышей лактация утрачивается) | Ormandy et al. (1997), Trott et al. (2012) |
| Поведение матери | Выхаживание потомства | Пролактин повышает материнскую ответственность перед потомством у крыс Пролактин изменяет поведение у Prlr -/- мышей | Rosenblatt (1967), Bridges et al. (1985, 1990, 1996), Bridges & Ronsheim (1990) Lucas et al. (1998) |
| Нейрогенез во взрослом возрасте | Повышенный нейрогенез в субвентрикулярной зоне материнского мозга | Запускается изменениями пролактина во время беременности Играет роль в изменениях настроения и поведения после родов | Shingo et al. (2003), Larsen et al. (2008), Larsen & Grattan (2010, 2012) |
| β-Клетки поджелудочной железы / гомеостаз | Материнские ткани становятся резистентными к инсулину, что способствует поступлению глюкозы к плоду Экспансия β-клеток матери для повышения продукции инсулина и предотвращения гестационного диабета | Рецепторы пролактина экспрессируются в β-клетках; экспрессия повышается во время беременности Пролактин стимулирует пролиферацию β-клеток и секрецию инсулина Нарушенная толерантность к глюкозе во время беременности у Prlr -/- мышей | Moldrup et al.(1993), Sorenson & Stout (1995), Brelje et al. (2004) Karnik et al. (2007) Huang et al. (2009), Kim et al. (2010), Rieck & Kaestner (2010), Schraenen et al. (2010), Zhang et al. (2010) |
| Регуляция аппетита | Повышенный аппетит и развитие Отложение жира во время беременности, мобилизация жира во время лактации | Пролактин стимулирует потребление пищи у небеременных животных Пролактин способствует развитию лептинорезистентности во время беременности Пролактин способствует повышению аппетита по время лактации | Gerardo-Gettens et al. (1989), Sauve & Woodside (1996, 2000), Naef & Woodside (2007) Augustine & Grattan (2008), Ladyman (2008), Ladyman et al. (2010) Woodside et al. (2012) |
| Костный и кальциевый гомеостаз | Повышенные захват кальция и мобилизация запасов кальция для роста скелета плода и продукции молока | Рецепторы пролактина на остеобластах и хондроцитах Пролактин стимулирует костный метаболизм Пролактин способствует всасыванию кальция в кишечнике | Charoenphandhu et al. (2010), Wongdee et al. (2011) |
| Размножение | Поддержание беременности (у грызунов) Утрата репродуктивного цикла во время беременности и лактации | У грызунов пролактин стимулирует функцию желтого тела, необходимую для поддержания беременности Гиперпролактинемия вызывает бесплодие Возможное / вероятное участие в лактационном бесплодии | Gibori & Richards (1978), Bachelot et al. (2009) Patel Bamigboye (2007) McNeilly (2001), Liu et al. (2014) |
| Реакция на стресс | Сниженный ответ на стресс на поздних сроках беременности и во время лактации для уменьшения воздействия глюкокортикоидов на плод | У самцов и небеременных самок пролактин действует как анксиолитик и снижает реакцию на стресс Возможная роль во время беременности и лактации | Shanks et al. (1999), Torner et al. (2001), Lonstein (2007), Brunton Russell (2008) Torner Neumann (2002), Slattery Neumann (2008) |
| Секреция окситоцина | Значительные изменения паттерна секреции, появление пиковых выбросов для облегчения родов и лактации | Рецепторы пролактина на окситоциновых нейронах; быстрые ингибирующие эффекты на активность Стимуляция экспрессии гена окситоцина | Kokay et al. (2006), Sapsford et al. (2011) Parker et al. (1991), Ghosh Sladek (1995) |
| Секреция пролактина | Измененная обратная связь для повышения секреции пролактина | Пролактин продолжает стимулировать TIDA-нейроны, но секреция дофамина снижается Повышение продукции энкефалина в TIDA | Grattan et al. (2008), Romano et al. (2013) Merchenthaler et al. (1994), Nahi & Arbogast (2003) |
Нейроэндокринный контроль секреции пролактина
Дофамин как пролактин-ингибирующий гормон
Ингибирующая регуляция секреции пролактина была продемонстрирована еще в 1950-х гг., однако найти ингибирующий гормон, опосредующий этот эффект, не удавалось. Все гормоны гипоталамуса, идентифицированные на тот момент, имели пептидную структуру, и предполагалось, что «пролактин-ингибирующий фактор» тоже должен быть пептидом. Первые подозрения ошибочности этой теории появились после того, как было обнаружено, что некоторые препараты, такие как резерпин, снижающие уровни эндогенных катехоламинов, индуцируют псевдобеременность у грызунов (Barraclough & Sawyer,1959), что указывает на повышение пролактина. Тем не менее предполагалось, что функциональная роль катехоламинов как нейротрансмиттеров, действующих в гипоталамусе, состоит в регуляции высвобождения гипоталамических гормонов (Kanematsu et al., 1963).
Основываясь на данных о наличии терминалей дофаминергических нервов в срединной возвышенности гипоталамуса (Fuxe, 1963), Мак-Леод предположил, что дофамин может высвобождаться в портальную систему гипофиза и таким образом действовать как гипоталамический гормон (в отличие его нейротрасмиттерной роли в других системах) (MacLeod et al., 1970). Он продемонстрировал, что дофаминергические агонисты эффективно подавляют секрецию пролактина in vivo и что дофамин может ингибировать секрецию пролактина в изолированных гипофизарных железах (MacLeod et al., 1970). Впоследствии дофамин был обнаружен в портальной крови гипофиза (Kamberi et al., 1970), и ряд исследователей установили, что вариабельность уровней пролактина в портальной крови отвечает за изменения секреции пролактина при различных физиологических состояниях (Ben-Jonathan et al., 1977, 1980; Gibbs & Neill, 1978; De Greef & Neill, 1979).
В 1990 г. Mansour et al. идентифицировали рецепторы дофамина на лактотрофных клетках передней доли гипофиза. Наблюдение гиперпролактинемии у мышей, не имеющих D2-рецепторов дофамина (Kelly et al., 1997; Saiardi et al., 1997), убедительно продемонстрировало ключевую роль дофамина в подавлении секреции эндогенного пролактина (рис. 1). Дофаминовые нейроны, контролирующие секрецию пролактина, расположены в дугообразном ядре гипоталамуса.
В зависимости от анатомического проецирования эти нейроны можно разделить на три субпопуляции: тубероинфундибулярные (TIDA), туберогипофизарные (THDA) и перивентрикулярные гипофизарные (PHDA) дофаминергические нейроны (Freeman et al., 2000). TIDA-нейроны берут начало в дорсомедиальном дугообразном ядре и проецируются во внешнюю зону срединной возвышенности (Bjorklund et al., 1973). Клетки двух других популяций локализуются более рострально, их проекции проходят в гипоталамо-гипофизарном тракте через срединную возвышенность к гипофизу. THDA-нейроны располагаются в ростральном дугообразном ядре и проецируются в промежуточную и невральную доли гипофиза (Fuze, 1994; Holzbauer & Racke, 1985).
PHDA-нейроны происходят из перивентрикулярного ядра, их аксоны заканчиваются в промежуточной доле (Goudreau et al., 1992). TIDA-нейроны вызывают классическую секрецию гормонов гипоталамуса в портальные сосуды гипофиза, а THDA- и PHDA-нейроны участвуют в базальной регуляции секреции пролактина после поступления дофамина в переднюю долю гипофиза по коротким портальным сосудам из нейрогипофиза (Peters et al., 1981). Несмотря на различное анатомическое расположение, три вышеуказанных типа нейронов имеют в значительной степени пересекающиеся дендритические поля (van den Pol et al., 1984) и одинаково регулируются. Например, все три типа нейронов стимулируются пролактином (DeMaria et al., 1999). Следовательно, их можно рассматривать как функциональную единицу пролактинингибирующих нейронов.
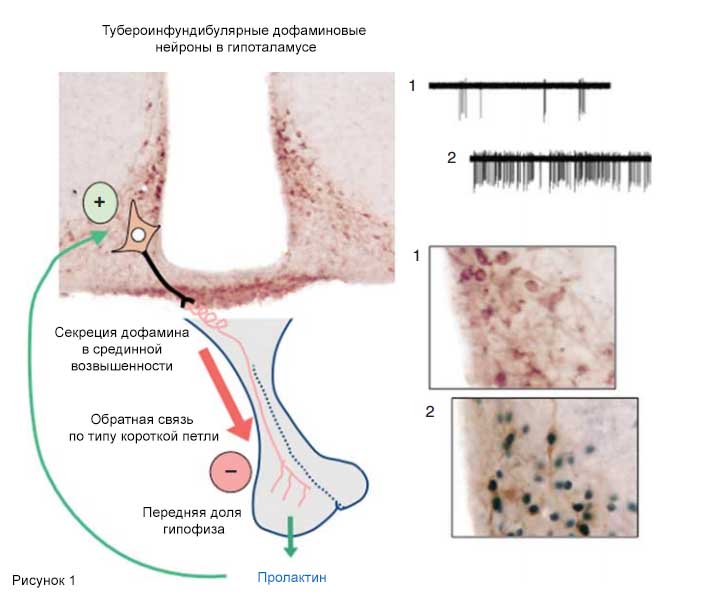
Рис. 1. Схематическое представление нейроэндокринной регуляции секреции пролактина.
Высвобождение пролактина передней долей гипофиза ингибируется дофамином, поступающим из тубероинфундибулярных дофаминовых нейронов (показаны на левой части рисунка; коронарная проекция, иммуногистохимическое исследование для выявления тирозингидроксилазы), тела которых располагаются в дугообразном ядре гипоталамуса, а аксоны проецируются в наружный слой срединной возвышенности. На правой части рисунка показаны примеры быстрой обратной связи (электрофизиологическая активация) и отсроченной обратной связи (фосфорилирование STAT5, черные ядра) в TIDA-нейронах; 1 - до назначения, 2 - после назначения пролактина.
В электрофизиологических исследованиях дофаминовых нейронов гипоталамуса у крыс TIDA-нейроны характеризовались четкой осцилляцией между гиперполяризованным и деполяризованным состояниями, с периодичностью около 20 с и частотой спонтанных разрядов примерно 4 Гц в деполяризованном рабочем состоянии. Примечательно, что TIDA-нейроны демонстрируют синхронный паттерн импульсации, т.е. они формируют единую сеть, работа которой зависит от фунциональной экспрессии щелевых контактов (Lyons et al., 2010). С помощью современных трансгенных технологий, позволяющих обозначить дофаминергические нейроны флуоресцентными метками, электрическая активность TIDA-нейронов также изучалась в срезах головного мозга мышей (Brown et al., 2012; Romano et al., 2013). При этом наблюдался такой же паттерн нейронных разрядов, как и у крыс, хотя в этой модели лишь небольшая часть TIDA-нейронов проявляла фазную осцилляцию. В одном из последних исследований с помощью амперометрии срединной возвышенности in vivo было продемонстрировано, что паттерны импульсации отдельных TIDA-нейронов являются отражением высвобождения дофамина (Romano et al., 2013). Эти данные поддерживают концепцию Lyons et al. (2010), согласно которой нейроны действуют как единая сеть, синхронно высвобождая дофамин в пульсирующем или фазном режиме.
Из пяти рецепторов дофамина в гипофизе были обнаружены два члена семейства D2-подобных рецепторов: D2 и D4 (Valerio et al.; 1994; Matsumoto et al., 1995). Воздействуя на эти рецепторы, дофамин ингибирует функцию лактотрофов (Mansour et al., 1990; Ben-Jonathan & Hnasko, 2001). Уникальность лактотрофов как клеток передней доли гипофиза состоит в том, что они обладают спонтанной электрической активностью в отсутствие гипоталамической стимуляции, при этом секрецию пролактина стимулирует инфлюкс Са2+ через вольтажзависимые кальциевые каналы (VGCC) (Gregerson, 2006). Это объясняет высокие уровни базальной секреции пролактина и согласуется с регуляторными механизмами, основанными преимущественно на ингибировании. Дофамин ингибирует инфлюкс кальция, вызывая гиперполяризацию клеточной мембраны (Gregerson et al., 1994; Gregerson, 2003) и снижение секреции пролактина (Lledo et al., 1990). Помимо непосредственного влияния на секрецию пролактина, дофамин подавляет активность аденилатциклазы, что снижает экспрессию гена пролактина (Maurer, 1982; Elsholtz et al., 1991; Ishida et al., 2007).
Дофамин также играет значительную роль в регуляции пролиферации лактотрофов, что было продемонстрировано в культурах клеток гипофиза (Ishida et al., 2007) и in vivo в отношении эстрадиол-индуцированной пролиферации (Borgundvaag et al., 1992). При повышении уровней дофамина, например при утрате дофаминового транспортера, возникает резкое постнатальное снижение пролиферации лактотрофов, что в свою очередь приводит к значительному уменьшению количества этих клеток в возрасте 8 нед (Bosse et al., 1997). Утрата D2-рецептора, напротив, вызывает выраженную гиперплазию лактотрофов (Kelly et al., 1997; Saiardi et al, 1997) с последующим развитием пролактином. Гиперплазия лактотрофов усугубляется с возрастом и чаще наблюдается у женщин (Saiardi et al., 1997, Asa et al., 1999).
Склонность к гиперплазии лактотрофов и ускоренное образование опухолей гипофиза у женщин также обусловлены прямыми эффектами эстрадиола, который стимулирует продукцию пролактина лактотрофами. У D2-нокаутированных мышей гонадэктомия уменьшает гиперплазию лактотрофов и формирование опухолей, однако заместительная терапия эстрадиолом после гонадэктомии не полностью восстанавливает исходные уровни гиперплазии, то есть в яичниках присутствуют и другие факторы, способствующие пролиферации лактотрофов (Hentges & Low, 2002).
Пролактиновая регуляция дофаминовых нейронов: обратная связь по типу короткой петли
Как уже указывалось, гипоталамо-пролактиновая система не имеет специфичной эндокринной мишени и, как следствие, классической гормонально-зависимой отрицательной обратной связи, описанной для всех остальных гормонов передней доли гипофиза. Тем не менее эта система все же регулируется отрицательной обратной связью: пролактин сам обеспечивает афферентный сигнал в процессе, известном как обратная связь по типу короткой петли. Присутствие рецепторов пролактина на дофаминовых нейронах (Lerant & Freeman, 1998; Grattan, 2001, Kokay & Grattan, 2005) предсказывалось в ранних нейрохимических исследованиях, в которых было установлено, что экзогенный пролактин стимулирует синтез (Hokfelt & Fuxe, 1972) и обмен (Eikenburg et al., 1977, Annunziato & Moore, 1978) дофамина в гипоталамусе, повышенный метаболизм дофамина в срединной возвышенности (Lookingland et al., 1987) и секрецию дофамина в портальную систему гипофиза (Gudelsky & Porter, 1979). В то же время гипопролактинемия, индуцируемая назначением агонистов дофамина, подавляет секрецию дофамина (Arbogast & Voogt, 1991), то есть базальная активность дофаминовых нейронов зависит от эндогенных уровней пролактина в крови. Используя эти биохимические показатели активности TIDA-нейронов, действие пролактина можно описать как двухфазное: быстрый компонент повышенной активности наблюдается через 2-4 ч после назначения пролактина (Selmanoff, 1985), а через 12 ч проявляется отсроченный компонент (Demarest et al., 1984, 1986).
Более современные электрофизиологические данные свидетельствуют о еще более быстром действии пролактина на TIDA-нейроны у мышей (Brown et al. 2012, Romano et al. 2013) и крыс (Lyons et al. 2012). Эти исследования показали, что пролактин уже через несколько секунд или минут после применения вызывает 4-кратное повышение частоты импульсации нейронов, быстро изменяя паттерн импульсации с базального фазного в тонически активный. Таким образом, существует множество механизмов пролактиновой регуляции этих нейронов, задействующихся в различные временные промежутки. Обратная связь пролактина резко нарушается у мышей, у которых отсутствует фактор транскрипции STAT5b (Grattan et al., 2001), вероятно, вследствие нарушения долгосрочной регуляции экспрессии тирозингидроксилазы - фермента, лимитирующего скорость синтеза дофамина (Arbogast & Voogt 1991, Ma et al., 2005). Этот эффект может отвечать за отсроченный компонент обратной связи пролактина, требующий синтеза белка (Johnston et al. 1980), однако маловероятно, что он участвует в более быстрых компонентах обратной связи по типу короткой петли.
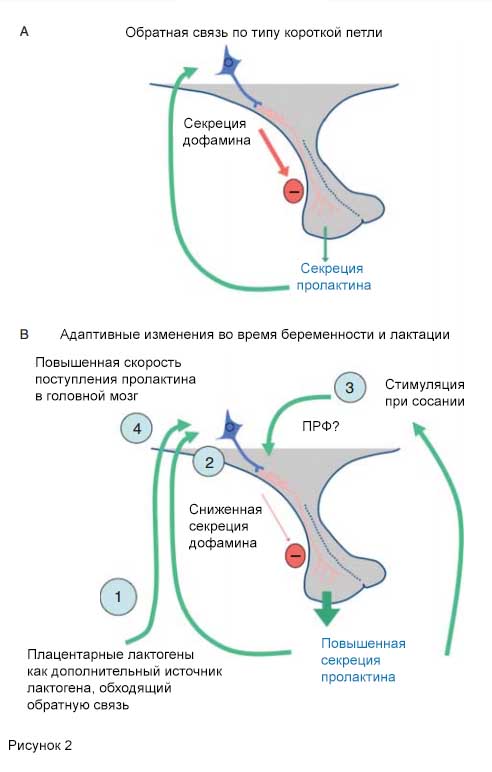
Рис. 2. А - схематическое изображение короткой петли обратной связи, контролирующей секрецию пролактина.
Б - адаптивные изменения в регуляции секреции пролактина во время беременности и лактации.
Существует несколько адаптивных механизмов, поддерживающих повышенные уровни лактогенных гормонов в крови и головном мозге матери и регулирующих широкий спектр функций для облегчения лактации: 1) продукция пролактиноподобных молекул в плаценте, что позволяет обойти регуляцию секреции пролактина в гипофизе по механизму обратной связи; 2) пластичность ответа TIDA-нейронов на пролактин: сниженная секреция дофамина и индукция экспресии экспрессии энкефалина; 3) адаптация поведения матери к сосанию, что создает наиболее мощный стимул высвобождения пролактина; 4) повышенное поступление пролактина в головной мозг во время лактации.
Очень быстрое действие пролактина, обнаруженное в электрофизиологических исследованиях, по-видимому, включает два компонента: низковольтажный, реализуемый TRP-током (током, подобным транзиторному рецепторному потенциалу), и высоковольтажный, связанный с Са2+-зависимым K+-током BK-типа. При этом высоковольтажный компонент является вортманнинчувствительным, что указывает на вовлечение сигнального пути PI3K (Lyons et al., 2012). В более медленном компоненте, который в первых нейрохимических экспериментах описывался как «быстрый» (от минут до часов), наиболее вероятно задействовано пролактин-индуцируемое серинфосфорилирование тирозингидроксилазы (Ma et al., 2005), повышающее активность фермента (Arbogast & Voogt, 1997). В совокупности эти три «слоя» пролактиновой регуляции дофаминовых нейронов гипоталамуса обеспечивают гомеостатический контроль, при котором пролактин быстро повышает частоту импульсации этих нейронов с последующим повышением секреции дофамина в портальную кровь и быстрым подавлением дальнейшей секреции пролактина лактотрофами. В то же время более медленные, но более продолжительные изменения в фосфорилировании тирозингидроксилазы и транскрипционных событиях, поддерживающие транскрипцию гена гидроксилазы, регулируют функцию нейронов в течение значительно более длительного периода, благодаря чему праймированные нейроны продолжают отвечать на изменения уровней пролактина (Grattan & LeTissier, 2015).
В отношении функции эндогенного пролактина в обратной связи по типу короткой петли, не совсем понятно, как этот относительно большой (197-199 аминокислот, 21 кДа) полипептидный гормон проходит через гематоэнцефалический барьер и достигает дофаминовых нейронов. Существует возможность, что дугообразное ядро и срединная возвышенность могут иметь неполный гематоэнцефалический барьер, и гормоны непосредственно попадают в нейроны этой области (Schaeffer et al., 2013), однако маловероятно, то пролактин посредством данного механизма регулирует дофаминовые нейроны гипоталамуса. Так, системное назначение пролактина одновременно активирует нейроны всего гипоталамуса (Brown et al., 2010; Sapsford et al., 2012), а не только дугообразного ядра. Существуют убедительные доказательства, что системный пролактин проникает через гематоэнцефалический барьер посредством насыщаемой транспортной системы с носителем (Walsh et al., 1987), в результате чего его уровни в цереброспинальной жидкости изменяются параллельно уровням в периферическом кровотоке (Login & MacLeod, 1977; Nicholson et al., 1980; Grattan & Averill, 1991). Учитывая высокую экспрессию рецепторов пролактина и его значительное связывание в хороидном сплетении (Walsh et al., 1978, 1990; Pi & Grattan, 1998; Augustine et al., 2003), предполагается, что рецепторы пролактина могут опосредовать его поступление в спинномозговую жидкость. В то же время недавно было продемонстрировано, что транспорт пролактина в головной мозг не зависит от пролактиновых рецепторов, поскольку он наблюдался у мышей, нокаутированных по рецепторам пролактина. Таким образом, точный механизм транслокации пролактина пролактина из кровяного русла в спинномозговую жидкость еще предстоит установить.
Grattan D.R. 60 Years of Neuroendocrinology: The hypothalamo-prolactin axis. J Endocrinol. 2015 Aug; 226(2): T101–T122.
Автор: Д.Р. Граттан, Центр нейроэндокринологии, Университет Отаго, Новая Зеландия